
Capítulo de livro publicado no livro do I CONGEB. Para acessa-lo clique aqui.
DOI: https://doi.org/10.53934/9786585062046-9
Este trabalho foi escrito por:
Ana Luíza Trajano Mangueira de Melo¹*; Cleidiane Macêdo Santos¹ ; Renata Ariel Fragoso Silva¹ ; Mariele Porto Carneiro Leão²;Antonio Félix da Costa²; José Ribamar Costa Ferreira Neto¹
¹Univeridade Federal de Pernambuco
² Instituto Agronômico de Pernambuco
*Ana Luíza Trajano Mangueira de Melo (Corresponding author) –[email protected]
As espécies vegetais são infectadas por vários microrganismos patogênicos que comprometem a sua sobrevivência e aptidão. Como consequência, ocorre redução significativa da produtividade anual das culturas. Em resposta a essa interação biológica nociva, as plantas empregam uma ampla gama de mecanismos de defesa, incluindo a síntese de membros do supergrupo das Pathogenesis-related proteins (PR’s), para evitar – ou minimizar – os danos ao seu metabolismo. As PR´s são componentes centrais da defesa das plantas a estresses bióticos e abióticos e compreendem 17 famílias (PR1-PR17). Entre elas, a PR-1 constitui a principal família de proteínas PR, sendo amplamente sintetizada em resposta ao ataque a uma gama de patógenos. Seus membros promovem o espessamento da parede celular vegetal e sequestram de esteróis da membrana dos patógenos, impedindo a sua proliferação. Muitos trabalhos têm abordado acerca da expressão família gênica PR1 e do seu caráter protetor contra estresses bióticos. No entanto, pouco se discute sobre a influência desses elementos na resposta de defesa de cultivares resistentes e suscetíveis, bem como no que concerne aos mecanismos que potencializem a sua indução durante processos infecciosos. Diante da importância da família PR1, o presente trabalho teve como objetivo fornecer, a partir de trabalhos publicados nos últimos cinco anos, uma visão geral da família Pathogenesis-related proteins 1 (PR1) e do seu envolvimento na defesa contra patógenos em diferentes espécies pertencentes ao clado Viridiplantae.
Palavras-chave: imunidade vegetal; patógenos; plantas
NTRODUÇÃO
As espécies vegetais são infectadas por vários microrganismos patogênicos que comprometem a sua sobrevivência e aptidão (1). Como consequência, ocorre redução significativa da produtividade anual das culturas, além de representar uma séria ameaça à segurança alimentar nos níveis doméstico, nacional e global (2). Em resposta a essa condição de estresse, as plantas empregam uma ampla gama de mecanismos de defesa, incluindo a síntese de membros do supergrupo das Pathogenesis-related proteins (PR’s) (3), para evitar ou minimizar os danos ao seu metabolismo.
As PR´s são componentes centrais da defesa vegetal a estresses bióticos e abióticos. Tais proteínas compreendem 17 famílias (PR1-PR17) que são classificadas com base em suas semelhanças de sequência, atividades enzimáticas e propriedades estruturais (4). A família PR1 constitui a principal família de proteínas PR, sendo sintetizadas amplamente em resposta ao ataque de vários patógenos e/ou indutores hormonais, como ácido salicílico (AS) e ácido jasmônico (5, 6). Seus membros promovem o espessamento da parede celular vegetal, impedindo o crescimento do patógeno ao bloquear sua entrada nas vias apoplásticas (7). Além disso, atuam no sequestro de esteróis da membrana dos patógenos, promovendo um efeito inibitório no crescimento dos agentes patogênicos, especialmente de organismos fúngicos (8).
Em soja, 20 genes GmPR1 (Glycine max PR1) foram regulados positivamente sob pelo menos um estresse biótico causado por fungos (Fusarium oxysporum, Fusarium virguliforme, Phialophora gregata e Phakopsora pachyrhizi), insetos (Aphis glycines), nematóides (Heterodera glycines, Rotylenchulus reniformis), oomiceto (Phytophthora sojae) e vírus (soybean mosaic virus) (9). Em outro estudo (10), foi observado em alho (Allium sativum L.) uma elevada expressão dos genes AsPR1 (Allium sativum PR1), resultando em maior inibição da infecção do fungo patogênico do gênero Fusarium. Resultados semelhantes foram encontrados (11) ao identificar o aumento transcricional de genes PR-1 em trigo (Triticum aestivum), associados ao maior controle da infecção por Fusarium equiseti.
Diante do importante papel molecular desempenhado pela família PR1 em vegetais submetidos a infecções por microrganismos patogênicos, o presente trabalho tem como objetivo fornecer uma visão geral da família Pathogenesis-related proteins 1 (PR1) e o seu envolvimento na resposta de defesa contra patógenos em diferentes espécies vegetais pertencentes à Viridiplantae, nos trabalhos publicados nos últimos cinco anos.
SELEÇÃO DE ESTUDOS
A presente revisão de literatura foi realizada durante os meses de julho e agosto de 2022, a partir da base de dados ‘PUBMED’, ‘Science Direct’ e ‘Scielo’. Tal ação abrangeu artigos em inglês, português e espanhol publicados nos últimos cinco anos em nível nacional e internacional. Para a realização da pesquisa, foram usados como palavras-chaves: ‘Pathogenesis related protein 1 and plants’, ‘Pathogenesis related-protein 1 and plants response’, ‘PR1 and plant pathogen response’, ‘PR1 and plant pathogen defense’. Os critérios de inclusão foram: estudos da família gênica Pathogenesis related-protein 1 associados aos mecanismos de defesa em Viridiplantae, estudos entre os anos de 2018 a 2022. Já os critérios de exclusão foram estudos não referentes à família Pathogenesis related-protein 1 em Viridiplantae, estudos não associados a estresses bióticos e revisões de literatura.
DESENVOLVIMENTO
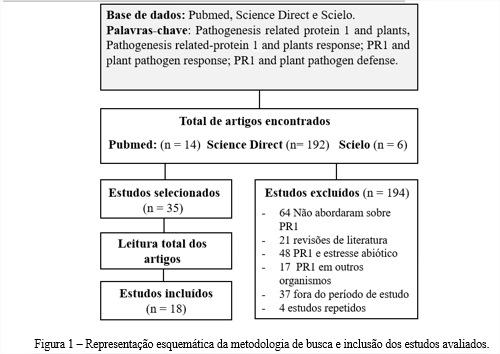
Ao total, por meio das três bases de dados utilizadas, foram encontrados 212 trabalhos durante a mineração de dados (Figura 1). Desses, 194 foram excluídos, dos quais: 64 não abordaram a temática das PR1; 21 se tratavam de revisões de literatura; 48 avaliaram PR1 associadas a estresses abióticos; 17 não avaliaram PR1 em Viridiplantae; 37 excederem a delimitação de período das publicações analisadas; e quatro estudos foram repetidos (Figura 1). Dezoito artigos, portanto, foram escrutinados na presente proposta.
ABORDAGENS DOS ESTUDOS ASSOCIADOS À FAMÍLIA PR1 EM VEGETAIS SOB ESTRESSES BIÓTICOS
A Tabela 1 apresenta os 18 artigos incluídos, os diferentes estresses bióticos aplicados nas plantas e os objetivos de cada estudo. Foi observado que as abordagens se dividiram entre a avaliação da expressão de PR1 em variedades de cultivares suscetíveis e resistentes submetidos à infecção por um ou mais fitopatógenos, além da análise da utilização de tecnologias voltadas para promover maior síntese de PR1 durante processos infecciosos.
Tabela 1 – Lista de artigos incluídos, estresses bióticos aplicados e objetivos.
| Autores | Estresse biótico | Objetivos |
| Almeida-Silva & Venancio (2022) (9) | Fusarium oxysporum, F. virguliforme, Aphis glycines, Heterodera glycines, Rotylenchulus reniformis e Phytophthora sojae | Caracterizar o gene PR-1 da soja (GmPR-1) nos níveis de sequência nucleotídica, estrutural e padrão transcricional em diversos tecidos sob condições de estresse biótico e abiótico. |
| Anuradha et al. (2022) (12) | Pseudocercospora eumusae e Pratylenchus coffeae | Analisar a família PR-1 de dois genomas da banana (Musa acuminata e M. balbisiana) e avaliar o padrão de expressão em ambas as espécies sob diferentes estresses. |
| Esmail, Aboulila & El-Moneim (2020) (13) | Puccinia striiformis | Compreender a variação na expressão dos genes PR1 e sua atividade enzimática após a inoculação com Puccinia striiformis em plântulas e adultos de milho (Triticum aestivum) . |
| Chun & Chandrasekaran (2019) (14) | Fusarium andiyazi | Avaliar o potencial das PR1 no controle da murcha causada por Fusarium andiyazi. |
| Aboulila (2022) (15) | Puccinia striiformis f. sp. tritici | Investigar a resistência sistêmica adquirida (SAR) induzida por cinco fitohormônios diferentes contra P. striiformis f. sp. tritici em variedades de trigo Egípcio e a sua associação com a expressão dos genes PR1 e PR4. |
| Butassi (2022) (16) | Taphrina deformans | Avaliar os mecanismos moleculares do pessegueiro (Prunus persica) em variedades suscetíveis e resistentes de plantas expostas a infecção pelo fungo Taphrina deformans. |
| Autores | Estresse biótico | Objetivos |
| Abbasi & Eslahi (2022) (17) | Rhizoctonia solani | Investigar o padrão de expressão de genes PR1 e a respectiva atividade enzimática em cultivares domesticados e selvagens de cevada (Hordeum vulgare) expostos à infecção por Rhizoctonia solani. |
Rabiei et al.(2022) (18) | Alternaria solani | Elucidar os efeitos do ácido hexanóico em variedades de tomates (S. lycorpesicum) submetidas à infecção por Alternaria solani. |
| Luo et al.(2022) (19) | Phytophthora infestans | Investigar os mecanismos moleculares associados à atividade de supressão de fungos oomycetos desempenhada por PR1 em batata (Solanum tuberosum). |
Dehghanian et al. (2020) (20) | Meloidogyne javanica | Examinar a indução de resistência ao tomateiro (S. lycopersicum) contra M. javanica e alterações na expressão de genes PR1, após aplicação de salicílico ácido e Pseudomonas fluorescens CHA0. |
| Farrakh, Wang & Chen (2018) (21) | Puccinia striiformis f. sp. tritici | Avaliar a expressão de genes PRs envolvidos em diferentes tipos de resistência do trigo à ferrugem. |
Jiang et al. (2022) (22) | Phytophthora capsici, Botrytis cinerea, Sclerotinia sclerotiorum e Erysiphe cucurbitacearum | Investigar a influência o óleo essencial de Litsea cubeba no desenvolvimento da SAR em diferentes espécies vegetais submetidas à infecção do vírus do mosaico do tabaco e doenças fúngicas. |
| Moghaddam et al. (2019) (23) | Alternaria alternata | Avaliar a resposta de sete genes PRs no genoma de diferentes variedades suscetíveis e resistentes de S. lycorpersicum durante a infecção por Alternaria alternata. |
| Liu et al. (2021) (24) | Alternaria alternata | Investigar o efeito da L-lisina exógena sobre o controle de A. alternata em frutos de pera e avaliar as mudanças de metabolismo de ROS e respostas de diferentes proteínas PRs |
| Autores | Estresse biótico | Objetivos |
| Wang et al. (2018) (25) | Phytophthora nicotianae eVírus do mosaico do tabaco (TMV) | Explorar a estratégia de indução de proteínas PRs em tabaco para combater Phytophthora nicotianae e TMV, com o auxílio de um extrato vegetal de plantas medicinais. |
Chaudhary, Singh & Charma (2021) (26) Gao et al. (2020) (27) | Ralstonia solanacearum Pseudomonas syringae | Avaliar in vitro a ação de ácido salicílico e ácido β-aminobutírico contra R. solanacearum e sua influência no padrão de expressão de genes PRs em cultivares suscetíveis e resistentes de tomate. Analisar a resistência de plantas de tabaco NtabSPL6-2 transgênicas durante a infecção por |
| Chu et al. (2022) (28) | Acidovorax avenae subsp. avenae | Avaliar os padrões de expressão de ScPR1 em resposta a estresses bióticos e abióticos em dois cultivares de cana-de-açúcar. |
Os vegetais avaliados nos estudos e pertencentes à Viridiplantae foram bastante diversificados, porém, majoritariamente de interesse econômico, como: tomate, banana, trigo, tabaco, soja, batata, cevada, cana-de-açúcar e pera. Entre os patógenos utilizados para a aplicação dos estresses bióticos, 17 (65%) foram fungos, quatro (15%) foram nematoides, três (11,5%) foram bactérias, um (3,8%) inseto e um (3,8%) vírus (Tabela 1).
AVALIAÇÃO DA EXPRESSÃO DE PR1 EM VARIEDADES DE DIFERENTES CULTIVARES SUSCETÍVEIS E RESISTENTES SUBMETIDOS A ESTRESSES BIÓTICOS
Como mencionado acima, muitos estudos aqui evidenciados avaliaram o perfil de expressão da família PR1 em variedades suscetíveis e resistentes frente à infecção por fitopatógenos (Tabela 1). Tal família destaca-se por ser transcricionalmente induzida por ácido salicílico durante processos infecciosos, compondo as vias responsáveis pelo desencadeamento da RSA (Resistência Sistêmica Adquirida) em plantas (29). Dessa forma, as PR1 consistem em uma família gênica de relevante interesse agronômico.
Dentre os estudos acima mencionados, Anuradha et al. (12) identificaram que as espécies resistentes de banana (Musa acuminata e M. balbisiana) apresentaram cinco genes da família PR1 com a expressão significativamente elevada, quando comparados ao grupo controle, em suas raízes durante a infecção pelo fungo F. oxysporum cubense. Em contrapartida, cultivares suscetíveis de ambas as espécies apresentaram expressão reduzida desses mesmos genes, sugerindo, dessa forma, a influência dos genes supracitados no processo de resistência a patógenos dessas plantas durante as interações incompatíveis com o referido fungo.
Nessa mesma linha, em outro trabalho (19), foram identificados 10 genes pertencentes à família PR1 em batata (S. tuberosum) dos quais foi possível verificar que as folhas da variedade resistente ‘E3’ apresentaram os 10 genes induzidos e com maiores níveis de expressão durante a infecção por P. infestans.A variedade suscetível ‘Desireé’, por sua vez, apresentou indução de oito desses genes. O estudo selecionou três isoformas (StPR1.2, StPR1.3 e StPR1.8) para avaliar o seu papel em plantas transgênicas. Foi observado que, nas plantas modificadas e com elevada indução de PR1, a resistência ao fungo foi significativamente maior, com o índice de doença (DI) inferior a 7. Nas variedades selvagens e no grupo e controle, por sua vez, os sintomas foram mais severos, com DI atingindo 80,4 e 80,8, respectivamente.
A expressão de PR1 em cultivares contrastantes em relação à resposta a patógenos também está presente em outro achado da literatura (16), no qual foi observado que PR1 foi induzido nas folhas tanto de variedades suscetíveis como de variedades resistentes de pessegueiro, após 96h de inoculação do fungo T. deformans. Uma maior elevação, ainda que sutil, dessa expressão foi relatada para a variedade resistente, porém sem significância estatística.
Sob outra perspectiva, ao avaliarem o papel desempenhado por genes da família PR1 em folhas de trigo submetidas à infecção por P. striiformis (21),foi verificado que dois desses elementos gênicos permaneceram induzidos do 1º ao 14º dia após a inoculação do fungo na variedade “high-temperature adult-plant (HTAP)”, durante a interação incompatível. Foi observado, adicionalmente, que ambos os genes apresentaram níveis de expressão significativamente maiores em interações incompatíveis quando comparados a interações compatíveis, sinalizando que esses elementos potencialmente influenciam no comportamento de resistência dessas variedades frente ao ataque de fitopatógenos fúngicos.
Em outro trabalho, que também utilizou cultivares de trigo (13), foi constatadoque os cultivares adultos e resistentes “Misr-3” e “Yr-29” apresentaram significativa indução dos genes PR1 após 72h de exposição a P. striiformis em comparação com o grupo controle. Tal fato contrastou com a expressão reduzida desses genes para o mesmo período em plântulas desses cultivares, após a inoculação fúngica. Além disso, foi observado que, nos cultivares suscetíveis “Sids-12” e “Avocet-S”, a expressão dos genes PR1 se mostrou induzida nas primeiras 24h, sendo reprimida no período após 72h.
Em conformidade com esses achados, ao avaliar o comportamento de cinco variedades suscetíveis e cinco variedades resistentes de tomate ao fungo A. alternata (23), foi observado que em cultivares adultos a expressão do gene PR1 foi elevada em todos os genótipos resistentes inoculados, com maior expressão observada nas variedades “Esfahan local”, “Rio Grande” e “H.a.s 2274”. Nas plantas suscetíveis, a indução do gene PR1 foi observada apenas nos cultivares adultos “Hedieh” e “Chief Falat”. Em contrapartida, no estágio de plântula o nível de expressão do gene PR1 foi significativamente induzido em todas as variedades resistentes infectadas mesmo após 20 dias de inoculação em comparação com o controle.
Em confronto os resultados supracitados e ainda utilizando variedades de tomate, foi desenvolvido um estudo (26) que, entre os achados obtidos, está a significativa indução de dois genes PR1(PR1a e PR1b) avaliados em folhas da variedade suscetível “Pusa Ruby” de tomateiro, enquanto esses mesmos genes foram reprimidos na variedade resistente “Hawaii7996”, ambos após 48h e 96h de exposição a R. solanacearum. Além disso, maior nível de indução dos genes foi relatado na variedade suscetível “Pusa Ruby”, tratada com 50 μM SA após 24 e 48h, tanto nas plantas expostas como nas não expostas a infecção por R. solanacearum.
Seguindo essa mesma linha, em um trabalho utilizando folhas da cultivar de cana-de-açúcar “ROC22” resistente ao fungo A. avenae subsp. Avenae (28), foi encontrada elevada indução ScPR1 24h após a inoculação, com um aumento 11 vezes maior de expressão em comparação ao controle (p < 0.01). Em contraste, nas folhas da cultivar “MT11–610”, suscetível ao agente biológico analisado, o nível máximo de expressão do gene foi observado em 48h, com 6,3 vezes aumento de expressão em relação ao controle. No geral, ScPR1 foi expresso mais rápido e em maior nível na cultivar resistente em contraposição a cultivar suscetível, após a inoculação com o patógeno.
Em uma abordagem diferente, ao avaliar o padrão de expressão de PR1 nas raízes de genótipos de cevada submetidos à infecção por R. solani (17),foi verificado que os cultivares domesticados “Yusuf” e “Jolgeh” apresentaram elevados níveis de indução do referido gene cinco dias após a infecção por R. solani quando comparados ao grupo controle (p <0.01) e aos cultivares selvagens TN374 e S09. Isso indicou que as variedades avaliadas de cevada domesticadas, possivelmente, apresentam maior poder de resposta a R. solani que suas contrapartes selvagens.
A partir do exposto, é possível inferir que PR1 potencialmente exerce proeminente influência no aspecto de resistência e suscetibilidade dos cultivares analisados, estando a sua maior indução gênica presente especialmente em variedades resistentes a partir de 24h após a exposição ao patógeno (28, 13, 21, 23), podendo se estender entre o 14º (21) até o 20º dia após a inoculação em algumas espécies (23). Dessa forma, essa família gênica se apresenta como elemento promissor para o desenvolvimento de tecnologias que possibilitem induzir a sua transcrição, de modo a promover respostas efetivas na defesa vegetal contra organismos patogênicos.
ESTRATÉGIAS PROMOTORAS DE MAIOR INDUÇÃO DE PR1 DURANTE PROCESSOS INFECCIOSOS
Como relatado, muitos estudos abordam a associação existente entre a indução da expressão gênica da família PR1 e a acentuada resposta de defesa vegetal contra fitopatógenos (5). No entanto, de acordo com a literatura (9), alguns agentes invasores são capazes de inibir a expressão da família gênica PR1. Entre os achados do referido estudo, foi encontrado que Phytophthora sojae e Fusairum virguliforme reprimiram sistematicamente a expressão dos genes PR-1 em soja (G. max), sugerindo que esses fungos podem regular negativamente a transcrição desses genes e influenciar no agravamento da doença. Dessa forma, alguns trabalhos foram desenvolvidos visando explorar potenciais meios de promoção da expressão de PR1 durante processos infecciosos. Entre esses, foi avaliado a expressão de PR1 em folhas de tomate-cereja (S. lycorpersicum) tratadas com ácido hexanóico (HA) e expostos à infecção por A. solani (22). Os autores observaram respectivos aumentos gradativos da expressão do gene em 24, 48 e 72h após a infecção. Além disso, foi constatado que a expressão de PR1 nos tratamentos inoculados com 1mM do HA após 72h de aplicação do fungo foi 57% maior quando comparado ao grupo controle e não tratado com o composto mencionado, havendo diferença significativa entre ambos.
Nessa mesma linha, outro grupo de pesquisa (14) objetivou investigar a expressão de genes PR1 induzidos por nanopartículas de quitosana e a sua implicação em cultivares de tomate-cereja (S. lycopersicum) submetidos a infecção por F. andiyazi (14). Nas raízes de tomate e quando comparado ao grupo controle (p<0.01), os autores constataram maior indução da expressão de PR1 mediada por nanopartículas de quitosana, 48h após a inoculação do patógeno. A elevação da expressão desses genes também foi significativamente maior quando comparado a inoculação exclusivamente com quitosana. Dessa forma, os resultados apontam para o potencial desses elementos enquanto nanopartículas influenciadoras do processo de resistência sistêmica adquirida da planta por meio da maior ativação de PR1.
De forma semelhante, foi observado que plântulas de tabaco (Nicotana glutnosa) apresentaram maior resistência aovírus do mosaico do tabaco (TMV) quando submetidaao tratamento com óleo essencial de citral (L. cubea) na concentração de 100 μg/m (22).O estudo utilizou como biomarcador molecular um gene PR1, identificando que em N. glutnosa tratada com 500 μg/mde citral houve elevado nível de indução desse gene nas suas folhas, 24h após a exposição ao TMV e a E. cucurbitacearum, B. cinerea e S. sclerotiorum. À vista disso, o tratamento com citral influenciou no desencadeamento da RSA da planta quando comparado ao controle (p<0.01).
Em conformidade com esses achados e também utilizando tabaco, foram avaliadas as estratégias de defesa da planta contra TMV e P. nicotianae (25). Os autores desse estudo observaram que após três aplicações da mistura de extratos das plantas S. flavescens e F. suspense combinada com agentes químicos, a expressão de PR-1 na espécie submetida à infecção por TMV e P. nicotianae aumentou em seis vezes quando comparados ao grupo controle. Ao serem utilizados separadamente, entretanto, tanto o extrato da planta quanto os agentes químicos induziram PR1 de forma menos eficaz. Dessa forma, o estudo demonstrou que o extrato foi capaz de induzir PR1 auxiliando no controle da infecção e apresentando como uma alternativa potencial ao controle das doenças em tabaco.
De forma semelhante, em outro trabalho, foi constatado que ao realizar o tratamento com lisina (0.1 mM) em folhas de pêra infectada por A. alternata, houve evidente redução da infecção pelo patógeno fúngico, sendo verificada a diminuição de 43.7 % no diâmetro da lesão após 15 dias de infecção (24). Os autores apontaram que um dos mecanismos moleculares que, potencialmente, foram envolvidos no controle da doença estiveram associados à maior indução de PR1 mediada pela lisina durante a infecção.
Buscando induzir a expressão de PR1 por meio de hormônios, um recente estudo (15) identificou genes PR1 com elevada indução nas folhas das variedades resistentes de trigo submetidas à infecção por P. striiformis e expostas a 1000 μg/m de ácido salicílico, especialmente a partir do 2º até o 7º dia após a infecção, sendo esse resultado significativo quando comparado ao grupo controle. Além disso, também foi constatada significativa e elevada expressão dos transcritos de PR1 nas folhas das variedades suscetíveis e resistentes de trigo [quando expostas a concentração de 150μg/ml ácido abscísico (ABA)], a partir do 1º dia após a infecção por P. striiformis.
Seguindo a mesma abordagem de exposição ao ácido salicílico, foi investigado o potencial de defesa de tomates contra M. javanica por meio da expressão de PR1, durante o tratamento com o mencionado ácido ePseudomonas fluorescens (rizobactéria que atua como bioagente contra patógenos do solo) (20). Foi evidenciado que as plantas expostas a M. javanica e inoculadas com ácido salicílico e P. fluorescens apresentaram maiores níveis de indução do gene PR1 após 48h de aplicação do estresse quando comparadas às plantas que foram tratadas exclusivamente com P. fluorescens ou ácido salicílico para o mesmo período. Tal diferença foi diretamente implicada na redução da infecção de M. javanica no tomateiro.
Sob outra perspectiva e utilizando uma abordagem diferente, foi identificado por meio de outro estudo (27), que o cultivar transgênico A. thaliana NtabSPL6-2 apresentou em suas folhas níveis significativamente maiores de indução de PR1 quando comparadas as plantas A. thaliana selvagem no 2º dia após a infecção por P. syringae. Os achados apontam PR1 como um biomarcador molecular indicativo da melhora da RSA mediada pelo gene inserido, NtabSPL6-2. Esse está envolvido no mecanismo de resposta defesa de Nicotina benthamiana contra patógenos.
Como evidenciado, os estudos recentes supracitados apresentaram como enfoque a investigação de meios alternativos que promovam maior expressão de PR1 durante o ataque de fitopatógenos em diferentes vegetais. Alguns achados apontam para resultados positivos utilizando estratégias como aplicação de ácido hexaônico (18), transgenia (27) ou nanopartículas de quitosana (14). Outra metodologia avaliada explorou formas de induzir a expressão de PR1 através da aplicação de diferentes concentrações de hormônios, como ácido salicílico (20, 15), o qual naturalmente induz a transcrição de PR1 m plantas durante estresses bióticos (4).
CONCLUSÕES
Em síntese, por meio do presente estudo, é possível inferir que a expressão dos genes pertencentes à família PR1 em vegetais exerce importante papel no controle de agentes infecciosos, especialmente organismos fúngicos que atingem Viridiplantae. Ademais, foi evidenciada forte associação entre a indução de PR1 e o seu o papel na resistência de cultivares pertencentes a diferentes espécies de interesse econômico, como tomate, banana, trigo, tabaco, batata e entre outros. Interessantemente, estudos demonstraram que a indução desses genes pode se iniciar 24h após a inoculação do patógeno e se estender até o 20º após a exposição ao estresse em alguns vegetais.
Outro aspecto observado diz respeito aos trabalhos encontrados na literatura que visam investigar meios de potencializar a indução de PR1 em plantas durante ataques por fitopatógenos. As metodologias empregadas para esse fim têm se mostrado eficientes e se dividem entre tecnologias com maior grau de complexidade, como o uso nanopartículas de quitosana e transgênicos e, com menor grau de complexidade, tal como a utilização do hormônio ácido salicílico, um indutor natural de PR1 em quadros infecciosos.
A presente revisão contribui para a maior compreensão sobre a importância do papel da família PR1 frente ao combate de organismos patogênicos, bem como para o seu potencial uso enquanto alvo a ser explorado por novas tecnologias voltadas para o melhoramento genético e que buscam ampliar a resistência de cultivares de interesse agronômico contra diversas doenças.
AGRADECIMENTOS
O presente trabalho foi desenvolvido com o apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) através da concessão da bolsa de Mestrado.
REFERÊNCIAS
- Savary S, Willocquet L, Pethybridge S J, Esker P, McRoberts N. The global burden of pathogens and pests on major food crops. Nat Ecol Evol. 2019; 3: 430–439.
- Fones HN, Bebber DP, Chaloner TM. Threats to global food security from emerging fungal and oomycete crop pathogens.Nat Food. 2020; 1: 455-456.
- Breen S, Williams SJ, Outram M, Kobe B, Solomon P S. Emerging Insights into the Functions of Pathogenesis-Related Protein 1. Trends in plant science. 2017; 22: 871–879.
- Ali S., Ganai BA, Kamili AN, Bhat AA, Mir ZA, Bhat JA. Pathogenesis-related proteins and peptides as promising tools for engineering plants with multiple stress tolerance. Microbiological research. 2018; 212: 29–37.
- Irigoyen M L, Garceau DC, Bohorquez-Chaux A. Genome-wide analyses of cassava Pathogenesis-related (PR) gene families reveal core transcriptome responses to whitefly infestation, salicylic acid and jasmonic acid. BMC Genomics. 2020; 21: 93-104.
- Jain D, Khurana J P. Role of Pathogenesis-Related (PR): Proteins in Plant Defense Mechanism and Molecular Aspects of Plant-Pathogen Interaction. Springer. 2018; 12: 124 – 136.
- Vandenberg N, Berger DK, Hein I, Birch PR, Wingfield MJ, Viljoen A. Tolerance in banana to Fusarium wilt is associated with early up-regulation of cell wall-strengthening genes in the roots. Mol Plant Pathol. 2007; 8 :333-41.
- Kazan K, Gardiner DM. Targeting pathogen sterols: Defence and counterdefence? PLoS Pathogology. 2012; 13.
- Almeida-Silva F, Venancio TM. Pathogenesis-related protein 1 (PR-1) genes in soybean: Genome-wide identification, structural analysis and expression profiling under multiple biotic and abiotic stresses. Gene. 2022; 809: 146-213.
- Anisimova, OK, Shchennikova, AV, Kochieva, EZ, Filyushin MA. Pathogenesis-Related Genes of PR1, PR2, PR4, and PR5 Families Are Involved in the Response to Fusarium Infection in Garlic (Allium sativum L.). Int. J. Mol. Sci. 2021; 22: 66-88.
- Manghwar H, Hussain A, Ali Q, Saleem MH, Abualreesh MH, Alatawi, A. Disease Severity, Resistance Analysis, and Expression Profiling of Pathogenesis-Related Protein Genes after the Inoculation of Fusarium equiseti in Wheat. Agronomy. 2021; 11, 21-34.
- Anuradha C, Chandrasekar A, Backiyarani S, Thangavelu R, Giribabu P, Uma S. Genome-wide analysis of pathogenesis-related protein 1 (PR-1) gene family from Musa spp. and its role in defense response during stresses. Gene. 2022;821:146334.
- Esmail SM, Aboulila AA, El-Moneim DA. Variation in several pathogenesis – Related (PR) protein genes in wheat (Triticum aestivum) involved in defense against Puccinia striiformis f. sp. tritici. Physiol Mol Plant Pathol. 2020;112:101545.
- Chun SC, Chandrasekaran M. Chitosan and chitosan nanoparticles induced expression of pathogenesis-related proteins genes enhances biotic stress tolerance in tomato. Int J Biol Macromol. 2019;125:948–54.
- Aboulila AA. Efficiency of plant growth regulators as inducers for improve systemic acquired resistance against stripe rust disease caused by Puccinia striiformis f. sp. tritici in wheat through up-regulation of PR-1 and PR-4 genes expression. Physiol Mol Plant Pathol. 2022;121:101882.
- Butassi E, Novello MA, Lara MV. Prunus persica apoplastic proteome analysis reveals candidate proteins involved in the resistance and defense against Taphrina deformans. J Plant Physiol. 2022;276:153780.
- Abbasi S, Eslahi N. Improved defense responses in modern barley cultivars compared to wild relatives against Rhizoctonia solani. Rhizosphere. 2022;21:100442.
- Rabiei Z, Hosseini S, Dehestani A, Pirdashti H, Beiki F. Exogenous hexanoic acid induced primary defense responses in tomato (Solanum lycopersicum L.) plants infected with Alternaria solani. Sci Hortic. 2022;295:110841.
- Luo X, Tian T, Feng L, Yang X, Li L, Tan X, et al. Pathogenesis-related protein 1 suppresses oomycete pathogen by targeting against AMPK kinase complex. J Adv Res [Internet]. 2022 [acesso em 19 de agosto de 2022]; Disponível em: https://www.sciencedirect.com/science/article/pii/S2090123222000339
- Dehghanian SZ, Abdollahi M, Charehgani H, Niazi A. Combined of salicylic acid and Pseudomonas fluorescens CHA0 on the expression of PR1 gene and control of Meloidogyne javanica in tomato. Biol Control. 2020;141:104134.
- Farrakh S, Wang M, Chen X. Pathogenesis-related protein genes involved in race-specific all-stage resistance and non-race specific high-temperature adult-plant resistance to Puccinia striiformis f. sp. tritici in wheat. J Integr Agric. 2018;17(11):2478–91.
- Jiang Y, Ji X, Zhang Y, Pan X, Yang Y, Li Y, et al. Citral induces plant systemic acquired resistance against tobacco mosaic virus and plant fungal diseases. Ind Crops Prod. 2022;183:114948.
- Moghaddam GA, Rezayatmand Z, Nasr Esfahani M, Khozaei M. Genetic defense analysis of tomatoes in response to early blight disease, Alternaria alternata. Plant Physiol Biochem. 2019;142:500–9.
- Liu Y, Li Y, Bi Y, Jiang Q, Mao R, Liu Z, et al. Induction of defense response against Alternaria rot in Zaosu pear fruit by exogenous L-lysine through regulating ROS metabolism and activating defense-related proteins. Postharvest Biol Technol. 2021;179:111567.
- Wang R, Wang S, Pan W, Li Q, Xia Z, Guan E, et al. Strategy of tobacco plant against black shank and tobacco mosaic virus infection via induction of PR-1, PR-4 and PR-5 proteins assisted by medicinal plant extracts. Physiol Mol Plant Pathol. 2018;101:127–45.
- Chaudhary G, Singh D, Sharma M. Effect of chemical elicitors on the differential expression pattern of PR genes in susceptible and resistant cultivars of tomato against bacterial wilt disease caused by Ralstonia solanacearum. Physiol Mol Plant Pathol. 2021;116:101689.
- Gao H, Zhang L, Zhang KL, Yang L, Ma YY, Xu ZQ. Tobacco NtabSPL6-2 can enhance local and systemic resistances of Arabidopsis thaliana to bacterial and fungal pathogens. J Plant Physiol. 2020;253:153263.
- Chu N, Zhou JR, Rott PC, Li J, Fu HY, Huang MT, et al. ScPR1 plays a positive role in the regulation of resistance to diverse stresses in sugarcane (Saccharum spp.) and Arabidopsis thaliana. Ind Crops Prod. 2022;180:114736.
- Barleen VP, Belkum AV, Summerbell RC, Crous, PW, Thomma BPHJ. Molecular mechanisms of pathogenicity: how do pathogenic microorganism develop crosskingdom host jumps?. FMS Microbiol. 2007; 31: 239-277.

{kind=link}