Agron Science
Capítulo de livro publicado no Livro da IV Mostra dos Trabalhos de Conclusão de Curso da Especialização em Vigilância Laboratorial em Saúde Pública. Para acessa-lo clique aqui.
doi.org/10.53934/9786599965821-06
Este trabalho foi escrito por:
Jacqueline Barbosa Mendonça de Oliveira1; Gabriela Bastos Cabral2
1 Pós-graduanda em Vigilância Laboratorial em Saúde Pública, Vigilância Epidemiológica, Instituto Adolfo Lutz – Centro de Laboratório Regional de Santos (SP); e-mail: [email protected]
2 Mestra em Biomedicina do Núcleo de Ciências Biomédicas, Instituto Adolfo Lutz – Centro de Laboratório Regional de Santos (SP)
Resumo: A Febre de Chikungunya se caracteriza como uma síndrome febril de início súbito, com artralgias intensas, evolução debilitante e possibilidade de cronificação. No Brasil a circulação do vírus do Chikungunya foi registrada em 2014 e atualmente está presente em todas as Unidade Federativas, sendo o país das Américas com o maior número de casos de dengue e Chikungunya. Em 2021 ocorreu um surto no estado de São Paulo (ESP), com 98% dos casos confirmados concentrados na Região Metropolitana da Baixada Santista (RMBS). Esse trabalho tem como objetivo evidenciar a flutuação de casos confirmados do vírus da Febre de Chikungunya no Brasil, com recorte para o estado de São Paulo. Os dados foram coletados em plataformas de domínio público on-line (DataSUS e CVE). Foram calculados e representados cartograficamente os coeficientes de incidência do agravo tanto para o país, quanto para o ESP. No período estudado o Brasil apresentou 627.149 casos confirmados, concentrando mais de 50% nos anos de 2017 e 2022. A região nordeste apresentou maior número de casos, seguida pela região sudeste. As características epidemiológicas predominantes foram “sexo feminino, faixa etária de 20 a 39 anos, e raça parda”. No período estudado o ESP apresentou um total de 18.262 casos confirmados, destes 80% concentrados no ano de surto, 2021. Dos dez municípios do ESP com maior número de casos (81,10% do total) cinco correspondem a municípios localizados na RMBS. O agravo se configura como um obstáculo para a saúde pública, permanecendo necessários investimentos na estrutura de vigilância sanitária e epidemiológica, na promoção em saúde, no saneamento básico e na educação.
Palavras–chave: Vírus Chikungunya, Mapeamento geográfico, Epidemiologia descritiva, Vigilância Epidemiológica, Saúde Pública.
INTRODUÇÃO
A Febre de Chikungunya (CHIK) se caracteriza como uma síndrome febril de início súbito, com artralgias intensas, evolução debilitante e possibilidade de cronificação (ROSS, 1956; MORRISON, 1979; SÃO PAULO, 2021).
Esse agravo é transmitido principalmente por mosquitos vetores do gênero Aedes sp., por isso é classificado como um arbovírus. Nas últimas décadas as arboviroses têm sido responsáveis por um elevado número de surtos, epidemias e casos de mortalidade pelo mundo (COSTA et al., 2019; MAEDA et al., 2022).
Como a alta densidade populacional é um dos fatores que propicia a inserção, transmissão e manutenção da circulação viral, e os vetores possuírem capacidade de
dispersarem diferentes vírus, existe a possibilidade de sobreposição de transmissão, ocorrendo assim uma co-circulação (NUNES et al., 2015; MORAIS et al., 2022).
No Brasil a circulação do vírus do Chikungunya (CHIKV) foi registrada em 2014, sendo caracterizada atualmente como endêmica, é uma das arboviroses de maior circulação territorial, juntamente com o dengue (DEN) (COSTA et al., 2019; YAKOB, 2022).
O Brasil atualmente é o país das Américas com o maior número de casos do DEN e CHIK (PAHO, 2022), se configurando como área de interesse de estudos de dispersão, flutuação e perfil epidemiológico.
Dentro do território nacional distinguimos as regiões nordeste e sudeste como as mais acometidas, sendo que a região sudeste é a mais populosa. Nesta região o estado de São Paulo (ESP) notificou apenas casos importados até 2015, mas a partir de 2016 casos autóctones também foram notificados. Em 2020 ocorreu um surto de CHIKV na unidade federativa (UF), e em 2021 ocorreu uma maior disseminação do agravo, com 98% dos casos confirmados se concentrando na Região Metropolitana da Baixada Santista (RMBS) (IBGE, 2012; SÃO PAULO, 2014, 2021; MAEDA et al., 2022).
Considerado como centro industrial do país, com o melhor Índice de Desenvolvimento Humano (IDH) nacional, e a presença do maior porto da América Latina (Porto de Santos) (IBGE, 2012; SÃO PAULO, 2013), o ESP se configura como área de interesse de estudos epidemiológicos sobre a CHIK.
Com o objetivo de evidenciar a flutuação dos casos, utilizando dados secundários de domínio público, esse trabalho se configura como retrospectivo, descritivo e quantitativo dos casos confirmados da Febre de Chikungunya no Brasil, com recorte para os casos confirmados no estado de São Paulo.
METODOLOGIA
Os casos confirmados de CHIK e as características epidemiológicas (raça, sexo e faixa etária) do país foram coletados na plataforma de domínio público DataSUS, através do aplicativo TabNet Win32 3.0 (BRASIL, 2022b), do ano de 2017 a 2021. Para o ano de 2022 foi considerado o BEPA (BRASIL, 2022a). Os casos confirmados de CHIK no ESP por município foram obtidos na plataforma de domínio público de dados estatísticos do CVE, do ano de 2015 a novembro de 2022 (SÃO PAULO, 2022).
Foram elaborados dois bancos de dados no software Microsoft Excel 2013, utilizados para avaliação descritiva por ano, UF, municípios, características epidemiológicas e coeficiente de incidência do agravo.
Os coeficientes de incidência foram calculados por UF (Brasil), e por município (ESP). Este coeficiente foi determinado através da razão entre o número de casos confirmados e a população estimada de determinada área geográfica (IBGE), sendo expresso por 100 mil habitantes (hab.) (BRASIL, 2021, 2022a). Os coeficientes foram apresentados utilizando o software Quantum GIS Desktop versão 3.16.10 (Projeção SIRGAS 2000 / EPSG:4674).
RESULTADOS E DISCUSSÃO
No período de 2017 a 2022, o Brasil apresentou um total de 627.149 casos confirmados para CHIK. Dois anos do período estudado concentraram mais de 50% dos casos (26,52% em 2017 e 27,04% em 2022). O segundo ano de maior número de casos nacionais foi 2017, corroborando com Faria et al. (2022). A região nordeste foi a que concentrou maior número de casos (61,22%), seguida pela região sudeste (29,87%) (Fig. 1).
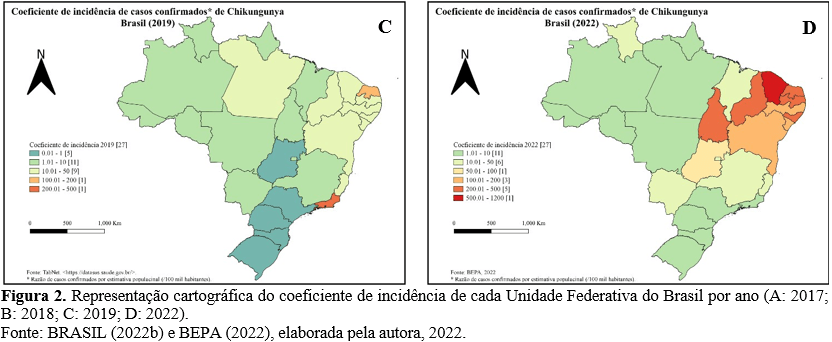
Após um pico em 2017 (129.385 casos) (Fig. 2.A), a região nordeste apresentou uma queda com manutenção da circulação do vírus até o ano de 2022 (Fig. 2.D), no qual volta a se configurar como região mais acometida (147.367 casos). De Oliveira et. al. (2021) inferiram que a redução no número de casos ao longo de 2018 – 2020 se deveu à diminuição de população suscetível. Além disso, é demonstrado um pico na região sudeste em 2019 (81.645 casos) (Fig. 2.C).
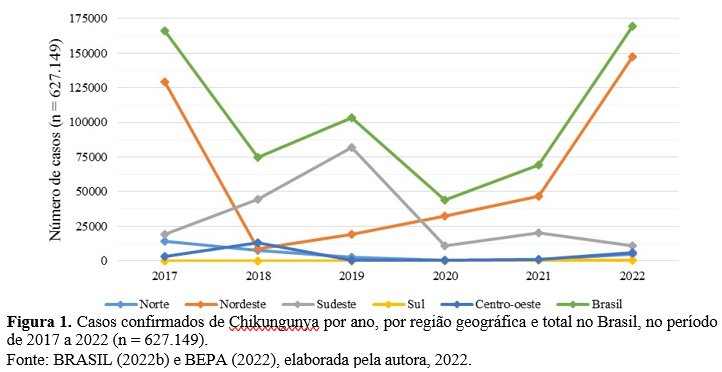
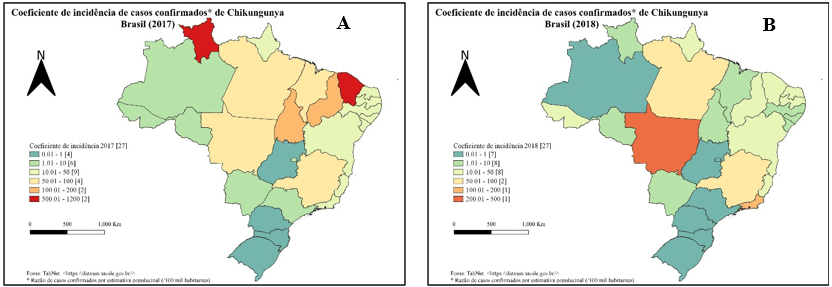
As UFs que apresentaram o maior número de casos durante o período estudado variaram entre as regiões nordeste e sudeste, corroborando com Da Silva Neto et al. (2022). Em cada ano do período estudado é possível identificar a UF mais acometida, sendo: Ceará em 2017 (64,89%) e 2022 (31,21%) (Fig. 2.A e 2.D), Rio de Janeiro em 2018 (43,51%) e 2019 (75,16%) (Fig. 2.B e 2.C), Bahia em 2020 (43,68%) e Pernambuco em 2021 (30,20%). Em 2021 o ESP foi o segundo estado com maior concentração de casos do país (21,46%).
Corroborando com Nunes et al. (2015), as regiões sudeste e nordeste são as que possuem maior potencial de estabelecimento do CHIKV. Os autores descreveram que 35 municípios do Brasil são importantes na manutenção da transmissão, sendo 57% localizados no nordeste e 31% no sudeste.
No ano de 2022 o BEPA (BRASIL, 2022a), até a SE 45, publicou que a região nordeste apresentou a maior incidência (255,5 casos / 100 mil hab.), seguida das regiões centro-oeste (35,8 casos / 100 mil hab.) e norte (25,7 casos / 100 mil hab.). Esse estudo obteve a região nordeste com maior incidência (281,98 casos / 100 mil hab.), seguida pela região norte (41,48 casos / 100 mil hab.) e pela região centro-oeste (28,14 casos / 100 mil hab.) (Fig. 2.D).
Em relação às características epidemiológicas as categorias predominantes nos casos confirmados no país foram: “parda” (51,30%), “feminino” (61,64%) e “entre 20 – 39 anos” (34,74%), corroborando com Nunes et al. (2015) e Silva et al. (2018).
A característica “raça” pode ser justificada pela composição étnica da região de maior incidência de casos. Segundo Costa et al. (2019) essa característica necessita ser analisada com cautela devido sua subjetividade. Além disso, sugerem que o número maior de casos entre as mulheres pode estar relacionado ao tempo de permanência em suas residências, sendo este o local de maiores focos do vetor, e por procurarem com mais frequência assistência médica. Indicam, também, que a “faixa etária” encontrada estaria relacionada à população economicamente ativa, que trabalha ou estuda durante o dia.
A caracterização epidemiológica encontrada neste estudo é verificada em outros países com registros de surtos e epidemias de CHIK (SISSOKO et al., 2009; SILVA et al., 2018).
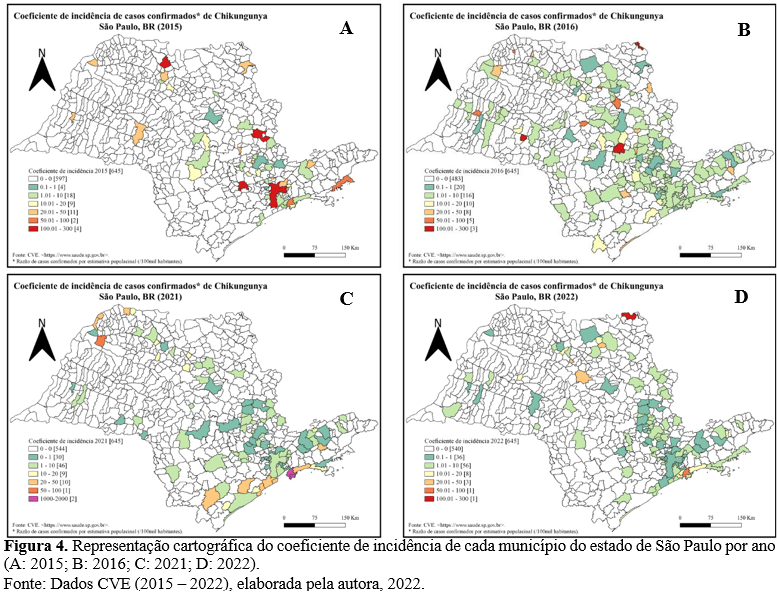
O ESP foi uma região de importante transmissão do CHIKV no ano de 2021, como a segunda UF de maior número de casos do ano (21,46%), sendo superado apenas por Pernambuco (30,20%). Ao longo do período estudado o ESP apresentou um total de 18.262 casos confirmados para CHIK (Fig. 3).
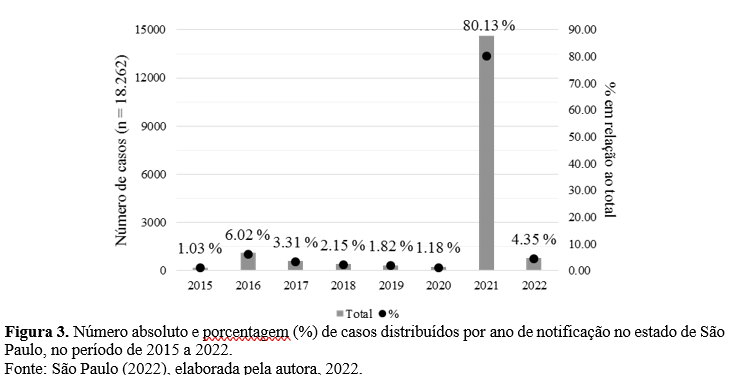
Durante o período de 2015 a 2020, no ESP os casos ocorreram de forma esporádica e com incidência entre 0,1 e 1,6 casos / 100 mil hab. (Fig. 4.A e 4.B). Em 2021 houve um crescimento exponencial, aumentando a taxa para 29,7 casos / 100 mil hab. (SÃO PAULO, 2021), correspondendo a 80,13% de todos os casos confirmados no estado no período estudado (Fig. 4.C).
No ano de 2021 ocorreu pela primeira vez transmissão expressiva do CHIKV no ESP, concentrando na RMBS 98% dos casos confirmados, com uma incidência de 715 casos
/ 100 mil hab. na região (SÃO PAULO, 2021; MAEDA et al., 2022). Esse fato é corroborado com os trabalhos de Silva et. al. (2018) e Costa et. al. (2019), que apresentaram a suscetibilidade da população como responsável pela transmissão efetiva (Fig. 4.C).
O ano com menor número de municípios com casos foi 2020 (32 municípios) e o maior foi 2016 (162 municípios). Até novembro de 2022 o ESP notificou 794 casos confirmados (105 municípios), sendo este o terceiro ano de maior número de casos no estado, depois de 2021 (14.634 casos) e 2016 (1.100 casos).
No ESP 10,26% (301 / 645) dos municípios apresentaram ao menos um caso de CHIK durante o período estudado (n = 18.262). Destes, 3,33% (10 / 301) dos municípios concentraram 89,74% dos casos (16.388). Dentre eles cinco municípios corresponderam à 81,10% de todos os casos e compõem a RMBS (Santos, Guarujá, São Vicente, Praia Grande e Cubatão). O município com maior representatividade de casos foi Santos (42,6%).
A flutuação dos casos no ESP obtida nesse estudo é corroborada pelas representações dos BEPA do CVE (SÃO PAULO, 2016, 2019 e 2020). Assim, é possível inferir que após a instalação do CHIKV na grande São Paulo e na RMBS houve manutenção da infecção nestas regiões, pois se apresentaram em destaque ao longo de todo o período.
CONSIDERAÇÕES
Após oitos anos de sua inserção no Brasil o vírus do Chikungunya apresentou capacidade de instalação e manutenção do ciclo de transmissão viral por todo o território, de forma que o país possui o maior número de casos absolutos nas Américas.
A partir dos coeficientes de incidência foi possível identificar o nordeste como região de maior incidência e o sul como região de menor incidência de casos confirmados. Assim como, foi possível identificar anos de transmissão expressivos em outras regiões.
Ainda que a circulação do vírus esteja presente no ESP desde 2015, existe a possibilidade de uma epidemia. Isso se deve ao fato de que, somente em 2021 houve uma transmissão expressiva no estado, concentrada na RMBS, e mais de 50% dos municípios do estado não apresentaram casos confirmados durante o período estudado.
As características epidemiológicas predominantes encontradas para os casos confirmados se mostram coerentes com outros estudos, porém a caracterização do perfil epidemiológico e de transmissão do Chikungunya no país se configura como complexa.
Desta forma, o agravo se configura como um obstáculo para a saúde pública, permanecendo necessários investimentos na estrutura de vigilância sanitária e epidemiológica, na promoção em saúde, no saneamento básico, na educação, e, não obstante, na pesquisa para que com dados atualizados e mais robustos o agravo possa ser melhor compreendido, monitorado, avaliado e controlado.
AGRADECIMENTOS
Agradeço à minha orientadora pela paciência, orientações, e confiança em meu conhecimento e produção. Agradeço às coordenadoras da regional pela disponibilidade, atenção e conhecimentos. Agradeço ao Instituto Adolfo Lutz pela oportunidade de vivenciar a experiência de pós-graduanda, absorvendo todos os conhecimentos que pude ter contato.
REFERÊNCIAS
ROSS, R. W. The Newala epidemic: III. The virus: isolation, pathogenic properties and relationship to the epidemic. Journal of Hygiene, 54(02), 177–191. 1956. DOI:10.1017/s0022172400044442.
MORRISON, J. C. L. Chikungunya Fever. International Journal of Dermatology, 18(8), 628–629. 1979. DOI:10.1111/j.1365-4362.1979.tb04677.x.
SÃO PAULO. Secretaria de Estado da Saúde de São Paulo. Coordenadoria de Controle de Doenças. Protocolo de Manejo Clínico de Chikungunya no Estado de São Paulo. 2021. 56p.
COSTA, A. K. S.; NINA, L. N. S.; CARVALHO, A. C.; BOMFIM, M. R. Q.; FELIPE, I.
M. A. Dengue e Chikungunya: Soroepidemiologia em usuários da Atenção Básica. Revista de Enfermagem UFPE On Line, v.13, n.4, p. 1006 – 1014. 2019. DOI: 10.5205/1981- 8963-v13i04a238828p1006-1014-2019.
MAEDA, A. Y. et al. Circulation of Chikungunya virus East-Central-South African genotype during the 2020–21 outbreak in São Paulo State, Brazil. Journal of Clinical Virology Plus, v. 2, n. 2, jun. 2022. DOI: 10.1016/j.jcvp.2022.100070.
NUNES, M. R. T. et al. Emergence and potential for spread of Chikungunya virus in Brazil.
BMC Medicine, v. 13, n. 1, abr. 2015. DOI 10.1186/s12916-015-0348-x
MORAIS, S. S. F. de et al. Mapeamento de Casos e Análise Inferencial da Incidência de Chikungunya em uma Capital do Nordeste Brasileiro. Archives Of Health Investigation, v. 11, n. 3, p. 403–410, mar. 2022. DOI: 10.21270/archi.vlli3.5709.
YAKOB, L. Predictable Chikungunya Infection Dynamics in Brazil. Viruses, v. 14, n. 9, ago. 2022. DOI: 10.339/v14091889.
PAHO. Pan American Health Organization. PLISA – Health Information Platform for the Americas. Chikungunya Cases. 2022.
SÃO PAULO. Secretaria de Estado da Saúde de São Paulo. Alerta 01 Surto de Chikungunya – estado de São Paulo: Sala de Situação Estadual de Enfrentamento às Arboviroses 2021. 2021.
IBGE. Instituto Brasileiro de Geografia e Estatística. Estimativas de População. Censo Brasileiro de 2010. Rio de Janeiro: IBGE. 2012.
SÃO PAULO. Centro de Vigilância Epidemiológica do Estado de São Paulo “Prof. Alexandre Vranjac”. Informe Epidemiológico – 09/06/2014, 2014
SÃO PAULO. São Paulo é o Estado com melhor IDH – Órgão da ONU divulgou nesta segunda o Índice de Desenvolvimento Humano dos municípios brasileiros. Do Portal do Governo, São Paulo, 29/07/2013. 2013
BRASIL. Ministério da Saúde. Banco de dados do Sistema Único de Saúde – DATASUS. TabNet. Epidemiológicas e Morbidade, Doenças e Agravos de Notificação, 2007 em diante (SINAN). Febre de Chikungunya. 2022b.
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Boletim Epidemiológico. Brasília, v.53, n.43, nov. 2022a.
SÃO PAULO. Centro de Vigilância Epidemiológica do Estado de São Paulo “Prof. Alexandre Vranjac”. Chikungunya – Dados Estatísticos, 2022.
BRASIL. Ministério da Saúde. Secretaria de Vigilância em Saúde. Departamento de Articulação Estratégica de Vigilância em Saúde. Guia de Vigilância em Saúde. 5ª ed. Brasília. 2021. 683 – 767. ISBN 978-65-5993-102-6.
FARIA, J. M. L. et al. Zika e Chikungunya entre 2017 e 2020: Um Estudo Epidemiológico. The Brazilian Journal of Infectious Diseases, v. 26, supl. 1, p. 120, jan. 2022. DOI: 10.1016/j.bjid.2021.101965.
DE OLIVEIRA, E. C. et al. Short report: Introduction of chikungunya virus ecsa genotype into the brazilian midwest and its dispersion through the americas. PLoS Neglected Tropical Diseases, v. 15, n. 4, abr. 2021. DOI: 10.1371/jornal.pntd.0009290.
DA SILVA NETO, S. R. et al. Arboviral disease record data – Dengue and Chikungunya, Brazil, 2013 – 2020. Scientific Data, v. 9, n. 1, mai. 2022. DOI: 10.1038/s41597-022-01312-
7.
SILVA, N. M. et al. Vigilância de chikungunya no Brasil: desafios no contexto da Saúde Pública. Epidemiologia e serviços de saúde: revista do Sistema Único de Saúde do Brasil, v. 27, n. 3, p. e2017127, set. 2018. doi: 10.5123/S1679-49742018000300003.
SISSOKO, D. et al. Post-Epidemic Chikungunya Disease on Reunion Island: Course of Rheumatic Manifestations and Associated Factors over a 15-Month Period. PLoS Neglected Tropical Diseases, v. 3, n. 3, mar. 2009. DOI: 10.1371/jornal.pntd.0000389.
SÃO PAULO. Secretaria de Estado da Saúde de São Paulo. Coordenadoria de Controle de Doenças. Boletim Epidemiológico Paulista. São Paulo, v. 13, n. 147, mar. 2016.
SÃO PAULO. Secretaria de Estado da Saúde de São Paulo. Coordenadoria de Controle de Doenças. Boletim Epidemiológico Paulista. São Paulo, v. 16, n. 188, agos. 2019.
SÃO PAULO. Secretaria de Estado da Saúde de São Paulo. Coordenadoria de Controle de Doenças. Boletim Epidemiológico Paulista. São Paulo, v. 17, n. 203, nov. 2020.
Capítulo de livro publicado no Livro da IV Mostra dos Trabalhos de Conclusão de Curso da Especialização em Vigilância Laboratorial em Saúde Pública. Para acessa-lo clique aqui.
doi.org/10.53934/9786599965821-05
Este trabalho foi escrito por:
Kelly Eduarda da Silva Sanches1; Bráulio Caetano Machado2; Rita de Cássia Compagnoli Carmona2
1Estudante do Curso de Especialização Adolfo Lutz – Núcleo de Doenças Entéricas/Centro de Virologia – Instituto Adolfo Lutz (NDE/CV-IAL); [email protected] 2Docente/pesquisador do NDE/CV – IAL; [email protected]
Resumo: A paralisia flácida aguda (PFA) é a apresentação clínica mais comum da infecção ocasionada pelo poliovírus 1, 2 e 3 (poliomielite). Em 1994 o Brasil recebeu da Organização Mundial de Saúde (OMS)/Organização Panamericana de Saúde (OPAS), a Certificação de área livre de circulação do Poliovírus Selvagem do seu território. À baixa taxa da cobertura vacinal e com a chegada de refugiados/repatriados de áreas com circulação do vírus, o Brasil corre risco de reintrodução do poliovírus. O objetivo deste estudo foi descrever os sorotipos de enterovírus não pólio (EVNP) a partir da vigilância dos poliovírus, fazendo a caracterização genotípica dos materiais de fezes de todo refugiado/repatriado positivo para EVNP, a fim de aumentar a sensibilidade da rede no Estado de São Paulo. As amostras de fezes foram sujeitas ao rastreamento de poliovírus pelo laboratório Instituto Oswaldo Cruz (IOC) – FIOCRUZ, a partir deste resultado, as amostras que deram positivas para ENVP foram submetidas à genotipagem pelo Núcleo de Doenças Entéricas, Centro de Virologia, Instituto Adolfo Lutz (NDE/CV/IAL) utilizando os métodos de purificação e suspensão fecal, transcrição reversa seguida da reação em cadeia pela polimerase convencional (RT- snPCR) para detecção do EVNP e reação de sequenciamento plataforma Sanger para determinação dos genótipos de EVNP. Foram recebidas pelo NDE/CV/IAL 340 amostras de fezes, os EVNP foram detectados em 5,3% (n=18/340) das amostras analisadas, destas 55,5% (N=10/18) foram detectadas e genotipadas, correspondendo ao CVA8 o tipo mais identificado, seguidos por E14, CVA19, E21 e EV-B84. Os dados obtidos contribuem para o conhecimento de quais EVNP estão circulando entre refugiados/repatriados durante janeiro a outubro de 2022 no Estado de São Paulo, Brasil.
Palavras–chave: enterovírus, enterovírus não pólio, paralisia flácida aguda, poliomielite, poliovírus
INTRODUÇÃO
Paralisia Flácida Aguda (PFA) é a manifestação clínica mais comum das infecções causadas pelo poliovírus 1, 2 e 3 (PV) conhecida como poliomielite. O vírus pertence ao
gênero Enterovirus (EV), família Picornaviridae, ordem Picornavirales. A família atualmente consiste em 158 espécies agrupadas em 68 gêneros. O gênero Enterovirus consiste em 15 espécies. Os EVs que infectam humanos são classificados em quatro espécies (EV-A, EV-B, EV-C e EV-D) (Tabela 1). Os sorotipos do poliovírus se encontram na espécie
C. Espécie EV-B tem sido descrito como principais agentes causadores de infecções do sistema nervoso (SNC) e meningite asséptica (MA). Entre esses tipos, os mais comuns são coxsackievirus B (CVB); coxsackievirus A9 (CVA9); echovirus (E) 4, 6, 9, 11, 13, 18, e 30 (CARMONA, R. de C.C, et al., 2020; HARVALA, H. et al., 2018; ICTV, 2020; RACANIELLO, V. R., 2006).
Após o controle dos PV os EVNP emergiram, causando surtos com manifestações clínicas muito similares à PFA, por isso o conhecimento desses vírus contribui para um diagnóstico diferencial, além de detectar e dar respostas a surtos.
OBJETIVO
O objetivo do trabalho é rastrear os enterovírus não pólio a partir da vigilância dos poliovírus, fazendo a caracterização genotípica dos materiais de fezes de todo refugiado/repatriado positivo para EVNP a fim de aumentar a sensibilidade da rede no Estado de São Paulo.
MATERIAL E MÉTODOS
Esse estudo foi realizado com 340 amostras fecais coletadas de refugiados/repatriados menores de 21 anos de idade, advindos de áreas de circulação de poliovírus, no período de janeiro a outubro de 2022.
A demanda de amostras foi estabelecida pela Nota Técnica Conjunta Nº 3/2022 (SES-SP/CCD/CVE/DDTHA/DVIMUNI/DVRESP/CIEVS-SP, 2022). As amostras fecais foram recebidas pelo Núcleo de Doenças Entéricas, Centro de Virologia, Instituto Adolfo Lutz (NDE/CV/IAL), Laboratório de Referência para os Enterovírus no estado de São Paulo e ponto focal da vigilância da PFA/Poliomielite, em seguida encaminhadas ao Laboratório de Referência Nacional das PFA/Poliomielite, Instituto Oswaldo Cruz (IOC), FIOCRUZ, Rio de Janeiro-RJ, para realização do diagnóstico laboratorial de poliovírus, conforme algoritmo preconizado pela OMS. Foram identificadas 18 amostras de fezes caracterizadas previamente como EVNP pelo IOC-FIOCRUZ, destas, 10 foram tipadas por caracterização molecular de genótipos de enterovírus pelo NDE/CV/IAL.
As amostras fecais estocadas a temperatura de -20°C foram preparadas de acordo com a metodologia descrita em Polio Laboratory Manual 2004 (WHO, 2004). Aproximadamente dois gramas de fezes foram adicionados a 10mL de uma solução tampão fosfato estéril, pH 7,4. As amostras foram agitadas e submetidas ao processo de centrifugação durante 60 minutos a 10.000xg em centrífuga refrigerada a 4°C. Em seguida, os sobrenadantes foram coletados e tratados com solução de penicilina (100U/mL) e estreptomicina (100ug/mL) por 30 minutos. Após este período os sobrenadantes foram estocados a baixas temperaturas -70° C até a realização dos testes.
O ssRNA dos EVNP foi extraído diretamente da suspensão fecal purificada, utilizando extrator automático Loccus, Kit – Extracta Fast 96 ou extração manual feita com o Kit Bio Gene ou Kit Qiamp viral RNA mini, de acordo com instruções do fabricante. A reação da transcrição reversa seguida da reação em cadeia pela polimerase convencional (RT-snPCR) foi realizada de acordo com o protocolo descrito por Nix, Oberste e Pallansch, (2006); WHO, (2015). A visualização dos produtos amplificados foi obtida por eletroforese em gel de agarose (1,5%) corado com corante de ácido nucleico (Gel Red® Nucleic Acid Stain – Biotium, INC., CA, USA). Foi utilizado marcador de peso molecular de 100pb (DNA Ladder, Invitrogen®). O sistema de corrida foi submetido a uma corrente elétrica constante de 100 Volts por 60 minutos. Após eletroforese os produtos amplificados foram observados em transiluminador de luminescência ultravioleta e o resultado captado pelo sistema de fotodocumentação digital UV Transilluminator, UVP Laboratory Products. A imagem salva foi impressa, analisada e arquivada.
Os produtos da RT-snPCR foram submetidos à reação de sequenciamento genômico pelo método de dideoxinucleotídeo, SANGER, F.; NICKLEN, S; COULSON, A.R. (1977), utilizando o kit BigDye® Terminator v3.1 Cycle Sequencing Ready Reaction Kit (Applied Biosystems®), com termociclagem de 25 ciclos de 96ºC por 10 segundos, 50°C por 5 segundos e 60°C por 4 minutos. Foram utilizados os mesmos primers da reação de RT- snPCR. O produto de sequenciamento foi precipitado utilizando o método de acetato e etanol. Os produtos precipitados foram ressuspendidos em 10μL de formamida Hi-Di® (Applied Biosystems®, Inc., USA) foram analisados no sequenciador automático modelo Applied Biosystems 3130xl Genetic Analyzer (Applied Biosystems®), utilizando o polímero POP-7® (Applied Biosystems®, Inc., USA).
As sequências dos genes obtidas foram editadas manualmente utilizando o programa Sequencher 4.7 (Gene Codes Corporation, Michigan, USA), para obtenção de sequências consensos. A ferramenta Enterovirus Genotyping Tool version 0.1 disponível no site http://www.rivm.nl/mpf/enterovirus/typingtool foi utilizada para confirmação dos genótipos de EVNP (Kroneman et al., 2011).
O TCC faz parte do Projeto de Pesquisa sob coordenação da Pesquisadora Científica Dra. Rita de Cássia Compagnoli Carmona, com aprovação quanto aos aspectos éticos pelo Comitê de Ética do Instituto Adolfo Lutz (IAL) e aprovação quanto ao mérito científico pelo Conselho Técnico Científico (CTC) do IAL – cadastro CTC 35-I /2016.
RESULTADOS E DISCUSSÃO
Entre os resultados positivos para EVNP disponibilizados pelo laboratório IOC- FIOCRUZ, 55,5% (n=10/18) foram caracterizados por sequenciamento genômico pelo NDE/CV/IAL (Tabela 1).
Os EVNP foram detectados em 5,3% (n=18/340) das amostras analisadas. 93,5% (n=318/340) negativas, n=4/340 (1,2%) poliovírus vacinal tipo 1 e 3, conforme representado no gráfico 1.
Dentre 5,3% (n=18/340) dos EVNP detectados, 55,5% (n=10/18) são genótipos caracterizados pelo NDE/CV/IAL. Cinco tipos distintos de EVNP foram identificados.
CVA8 espécie EV-A, foi o tipo mais comumente identificado (60%, n=6/10) seguido por E14 espécie EV-B, CVA19 espécie EV-C, E21 espécie EV-B, e EV-B84 espécie EV-B, cada um (10%, n=1/10) de acordo com o gráfico 2.
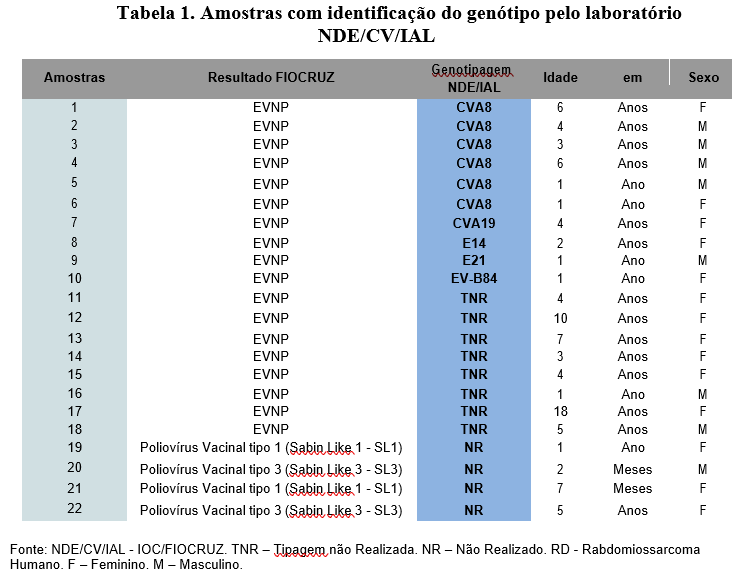
Gráfico 1 – Amostras de refugiados/repatriados menores de 21 anos entre janeiro a outubro de 2022
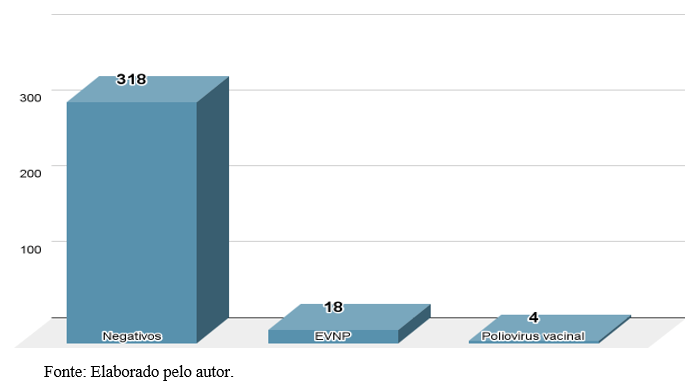
Gráfico 2 – Genótipos de EVNP caracterizados por sequenciamento genômico.
Os resultados obtidos neste estudo foram coletados de um grupo que não possui sintomas, com o objetivo de rastrear quais sorotipos de EVNP estão em circulação, diante da presença de pessoas advindas de área de circulação de poliovírus. É importante fazer o rastreamento dos EVNP, devido ao aumento dos casos de PFA ocasionados por esses vírus, relatando que nessa população estudada está ocorrendo à circulação de EVNP.
Diante da vigilância dos poliovírus no Brasil, os materiais coletados foram fezes, e por serem pessoas assintomáticas, que não possuem manifestações clínicas, não foi necessária a coleta de outro tipo de material biológico mais invasivo. Em casos de infecções neurológicas, as amostras de fezes e respiratórias têm sido consideradas as mais convenientes para serem submetidas ao teste de EV, além de LCR e sangue (CARMONA, R. de C.C, et al, 2020).
O coxsackievirus e echovirus foram relacionados a surtos com comprometimento motor (DIAS, A. P. M., 2018). O coxsackievirus A8 (CVA8), pertence à espécie EV-A foi o mais encontrado neste estudo, tem um perfil de causar doença febril, doença de mão pé boca, herpangina (HA), mas ocasionalmente levam a distúrbios neurológicos graves, como PFA. Em 2012 na Tailândia, o CVA8 foi a causa mais prevalente de HA. (SONG, Y. et al., 2020). O coxsackievirus A19 (CVA19) pertence à espécie EV-C, e tem sido associado a casos de doenças respiratórias e gastrointestinais, meningite asséptica, gastroenterite, PFA e enterite aguda (Yi, L., Zhang, L., Feng, L. et al, 2022).
O enterovirus B84 (EV-B84) pertence à espécie EV-B, causa viral mais comum de miocardite aguda, paralisia flácida aguda e meningite asséptica. A taxa de isolamento do vírus é extremamente baixa e não é um sorotipo prevalente no mundo. O primeiro relato do vírus na China foi de um paciente com PFA em 2004 (ZHENG, H. et al., 2016).
O echovirus 14 (E14) pertence à espécie EV-B está associado a casos de síndrome gastrointestinal, meningite asséptica, paralisia, necrose hepática neonatal fatal, o genótipo também é considerado raro (CHEN, P. et al., 2017). O echovirus 21 (E21) pertence à espécie EV-B, este genótipo não possui muitos isolados, mas já foi isolado de um paciente com PFA (LIU, H. et al., 2020).
No Brasil, um estudo de 1.215 amostras de paciente com suspeita de doenças infecciosas do SNC, realizado no Estado de São Paulo entre 2004 a 2014 por CARMONA, R. de C.C, et al (2020), identificou o E14 que foi associado a doença meningite asséptica (MA), e o E21 associado a meningoencefalite, ambos encontrados em (0,6%; n = 1/176) amostras positivas. E-6 foi o mais comumente identificado, seguido E-30; E18; CV-B5; E4; E11; CVB2 e E9; E7; CVA9, CVB1, CVB3, CVB4, E13, E14 e E21. Segundo CARMONA, R. de C.C, et al (2020) no Brasil os EVs foram detectados durante todo o ano e a variação sazonal é mais proeminente durante a primavera, já o pico de ocorrência foi no verão.
As amostras são de refugiados, sendo assim, esses genótipos podem ser comuns em seu país de origem, a chegada ao Brasil pode aumentar a probabilidade da introdução de genótipos não circulantes.
CONCLUSÕES
Os resultados encontrados de EVNP (CVA8, CVA19 E14, E21 e EV-B84), demonstram conhecimento de quais genótipos estão circulando entre refugiados/repatriados assintomáticos durante janeiro a outubro de 2022 no Estado de São Paulo, Brasil, demonstrando o perfil de circulação silenciosa dos EVs, que podem sofrer mutações até causarem manifestações clínicas mais graves, e em grandes surtos essas doenças podem ter uma grande proporção. Os achados não foram de casos de PFA, mas estudos anteriores relacionaram esses genótipos a casos de PFA. A vigilância epidemiológica e laboratorial tem feito um papel de extrema importância no controle e no rastreamento dos EVNP, a partir da grande comoção que a poliomielite tem causado com o risco de reintrodução no país, porém, vemos que outros agentes estão circulando com potencial de causar PFA e a probabilidade da introdução de novos genótipos no Brasil. Outros estudos precisam ser feitos para que os EVNP não venham causar grandes surtos semelhantes aos poliovírus.
AGRADECIMENTOS
Agradeço aos colegas Adriana Jorge, Amanda Dias, Antônio Erculiani, Daniele de Moraes, Fabricio Caldeiras, Gabriel Ferreira, Gisele Nemer e Lucas Leme do Núcleo de Doenças Entéricas, Centro de Virologia, Instituto Adolfo Lutz (IAL), que realizaram o diagnóstico dos Enterovírus Não Pólio, a Divisão de Doenças e Transmissão Hídrica e Alimentar, Centro de Vigilância Epidemiológica, Coordenadoria de Controle de Doenças, Secretaria de Estado da Saúde, São Paulo e o Laboratório de Enterovírus, Instituto Oswaldo Cruz (IOC), FIOCRUZ, Ministério da Saúde.
REFERÊNCIAS
CARMONA, R. de C.C, et al. Distribution of species enterovirus B in patients with central nervous system infections in São Paulo State, Brazil. Journal of Medical Virology. v. 92, ed. 12, 2020. Disponível em: <https://doi.org/10.1002/jmv.26131>. Acesso em: 02 nov. 2022.
HARVALA, H. et al. Recommendations for enterovirus diagnostics and characterisation within and beyond Europe. In: Journal of Clinical Virology. v. 101, p. 11-17, abr. 2018. Disponível em:
<https://doi.org/10.1016/j.jcv.2018.01.008>. Acesso em: 22 nov. 2022.
INTERNATIONAL COMMITTEE ON TAXONOMY OF VIRUSES EXECUTIVE COMMITTEE.
The new scope of virus taxonomy: partitioning the virosphere into 15 hierarchical ranks. Nat Microbiol. v. 5, ed. 5, p. 668-674, 2020. Disponível em: <doi: 10.1038/s41564-020-0709-x>. Acesso
em: 10 nov. 2022.
RACANIELLO, V. R. One hundred years of poliovirus pathogenesis. In: Virology. v. 344, ed. 1. 2006. Disponível em: <https://doi.org/10.1016/j.virol.2005.09.015>. Acesso em: 05 nov. 2022 SANGER, F.; NICKLEN, S; COULSON, A.R. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci U S A. ed.12, v. 74, p.5463-7. 1977.
SECRETARIA DE ESTADO DA SAÚDE DE SÃO PAULO. COORDENADORIA DE CONTROLE DE DOENÇAS – CCD. CENTRO DE VIGIL NCIA EPIDEMIOLÓGICA “PROF. ALEXANDRE VRANJAC”. DIVISÃO DE DOENÇAS DE TRANSMISSÃO HÍDRICA E ALIMENTAR – DDTHA. DIVISÃO DE IMUNIZAÇÃO – DVIMUNI. DIVISÃO DE DOENÇAS DE TRANSMISSÃO RESPIRATÓRIA – DVRESP. CENTRO DE INFORMAÇÕES ESTRATÉGICAS EM VIGIL NCIA EM SAÚDE –CIEVS-SP. NOTA CONJUNTA Nº 3/2022 –
DDTHA/CIEVS-SP/DVIMUNI/DVRESP/CVE/CCD/SES-SP. Alerta para risco de reintrodução da poliomielite no Brasil e recomendações para ações de Vigilância Epidemiológica das Paralisias Flácidas Agudas (PFA) junto aos refugiados/repatriados advindos de áreas de circulação de poliovírus (selvagem e/ou derivado vacinal). São Paulo, 2022.
WORLD HEALTH ORGANIZATION. Polio laboratory manual 4th edition.
Geneva 27, Switzerland: Department of Immunization, Vaccines and Biologicals CH- 1211, World Health Organization; p. 1–167. 2004.
NIX, W.A.; OBERSTE, M.S; PALLANSCH, M.A. Sensitive, seminested PCR amplification of VP1 sequences for direct identification of all enterovirus serotypes from original clinical specimens. J Clin Microbiol. v. 44, p. 2698–2704, 2006.
WORLD HEALTH ORGANIZATION. Enterovirus surveillance guidelines: Guidelines for enterovirus surveillance in support of the Polio Eradication Initiative, Geneva. World Health Organization; 2015. p.1-46.
SANGER, F.; NICKLEN, S; COULSON, A.R. DNA sequencing with chain-terminating inhibitors. Proc Natl Acad Sci U S A. ed.12, v. 74, p.5463-7. 1977.
KRONEMAN, A. et al. An automated genotyping tool for enteroviruses and noroviruses. Journal of Clinical Virology. v. 51, ed. 2, p. 121-5. jun. 2011. Disponível em: . Acesso em: 25 nov. 2022.
DIAS, A. P. M. Avaliação de um protocolo visando o diagnóstico rápido dos enterovírus associados a casos de paralisia flácida aguda. 2008. 73 f. Dissertação (Mestrado em Vigilância Sanitária) – Instituto Nacional de Controle de Qualidade em Saúde, Fundação Oswaldo Cruz, Rio de Janeiro, 2008.
SONG, Y. et al. Genetic Diversity Analysis of Coxsackievirus A8 Circulating in China and Worldwide Reveals a Highly Divergent Genotype. Viruses. v. 12, ed.10, p.1061, set. 2020. Disponível em: <https://www.mdpi.com/1999-4915/12/10/1061> . Acesso em: 25 dez. 2022.
ZHENG, H. et al. Isolation and Characterization of a Highly Mutated Chinese Isolate of Enterovirus B84 from a Patient with Acute Flaccid Paralysis. Sci Rep. v. 6, n. 31059, ago. 2016. Disponível em:
<https://www.nature.com/articles/srep31059> . Acesso em: 26 nov. 2022.
CHEN, P., et al. Filogeografia evolutiva e padrão de transmissão do echovirus 14: uma exploração da dinâmica espaço-temporal com base na vigilância da paralisia flácida aguda de 26 anos em Shandong, China. BMC Genomics. v. 18, n. 48, jan. 2017. Disponível em:<https://doi.org/10.1186/s12864-016-3418-3>. Acesso em: 27 nov. 2022.
LIU, H. et al. Caracterização de uma nova cepa de echovirus 21 isolada de uma criança saudável na China em 2013. Arch Virol. v. 165, p. 757–760, jan. 2020. Disponível em: <https://doi.org/10.1007/s00705-019-04506-4> Acesso em: 25 nov. 2022.
Capítulo de livro publicado no Livro da IV Mostra dos Trabalhos de Conclusão de Curso da Especialização em Vigilância Laboratorial em Saúde Pública. Para acessa-lo clique aqui.
doi.org/10.53934/9786599965821-04
Este trabalho foi escrito por:
Maria Marciana Pereira Silva¹; Tatiana Caldas Pereira²
1Estudante do Curso de Especialização em Vigilância Laboratorial em Saúde Pública – IAL; E-mail: [email protected]
2Docente/Pesquisadora do Núcleo de Ciências Químicas e Bromatológicas do Centro de Laboratório Regional de Santos do Instituto Adolfo Lutz – IAL; E-mail: [email protected]
Resumo: As Doenças de Transmissão Hídrica e Alimentar (DTHA) ocorre pelo consumo de água e/ ou alimentos contaminados por microrganismos patogênicos ou substâncias químicas. De acordo com o Ministério da Saúde existem mais de 250 tipos de DTHA que podem ser causadas pelo consumo de água e alimentos contaminados por bactérias, vírus e protozoários. A incidência de surtos de DTHA vem aumentando em diversos países, impactando em morbidade, mortalidade e custos econômicos. O Centers For Disease Control Prevention (CDC) dos Estados Unidos (EUA) estimou que os patógenos conhecidos de origem alimentar são responsáveis por 9,4 milhões de casos de DTHA todos os anos nos EUA, entre eles, o norovírus e a Salmonella spp. Na União Europeia (UE) o norovírus também é o principal agente etiológico envolvido em surtos de DTHA, assim como no Japão. Diferente dos EUA, UE e Japão, no Brasil e na Coréia, a bactéria E. coli é responsável pelo o maior número de surtos de DTHA. Conclui-se que os principais agentes etiológicos envolvidos em surtos de DTHA em diferentes regiões do mundo são norovírus, Salmonella spp, E. coli, Campylobacter spp, Staphylococcus aureus e Listeria monocytogenes.
Palavras Chave: Doenças Transmitidas por Alimentos, intoxicação alimentar, zoonoses, segurança alimentar, saúde pública.
INTRODUÇÃO
As Doenças de Transmissão Hídrica e Alimentar (DTHA) ocorrem pelo consumo de água e/ ou alimentos contaminados por microrganismos patogênicos ou substâncias químicas. De acordo com o Ministério da Saúde existem mais de 250 tipos de DTHA que podem ser causadas pelo consumo de alimentos contaminados por bactérias, vírus e protozoários (BRASIL, 2022).
A incidência de surtos de DTHA vem aumentando em diversos países, impactando em morbidade, mortalidade e custos econômicos. Considera-se um surto de DTHA quando duas ou mais pessoas apresentam doença ou sintomas semelhantes após ingerir alimentos contaminados. Para agravos como Cólera, Botulismo e Febre Tifoide a confirmação de apenas um caso já é considerado surto (NYACHUBA, 2010; BRASIL,2022).
Com base no relatório publicado em 2015 pela Organização Mundial da Saúde (OMS), aproximadamente 600 milhões de casos de DTHA foram registrados no ano de 2010 em todo o mundo, e cerca de 350 milhões das notificações estavam relacionadas a ingestão de alimentos e água contaminados por bactérias patogênicas (WHO, 2015).
Vários fatores contribuem para o aumento de surtos de DTHA, entre os quais, se destacam o aumento da população em geral, a existência de grupos populacionais vulneráveis ou mais expostos, o aumento da urbanização desordenada, a necessidade de produção de alimentos em larga escala, a globalização e o comercialização de produtos alimentícios entre países (ANTUNES et al.,2020).
Os surtos epidemiológicos associados às DTHA são um problema de Saúde Pública que atinge tanto os países desenvolvidos como os países em desenvolvimento. Cada país possui legislações específicas para o monitoramento e controle das Doenças de Transmissão Hídrica e Alimentar. Apesar dos esforços para conter as DTHA, o número de surtos vem crescendo nos últimos anos, afetando diretamente a saúde da população, sobrecarregando os sistemas públicos de saúde, gerando altos custos e impactando diretamente na economia (WHO, 2015). Desta forma, o presente estudo teve como objetivo verificar os principais agentes etiológicos envolvidos em casos e surtos de DTHA em diferentes regiões do mundo.
METODOLOGIA
Trata-se de um estudo de revisão bibliográfica relacionada a surtos de DTHA no Brasil e em diferentes regiões do mundo. Foram consultados artigos científicos nas bases de dados Scielo e Pubmed publicados no período de 2009 a 2022.
Para a busca dos artigos, foram utilizados os descritores: Doenças Transmitidas por Alimentos, intoxicação alimentar, zoonoses, segurança alimentar e saúde pública.
RESULTADOS E DISCUSSÃO
Nos Estados Unidos, o Centers For Disease Control Prevention (CDC) usou dados da vigilância ativa e passiva e outras fontes para estimar que patógenos conhecidos de origem alimentar são responsáveis por 9,4 milhões de casos de DTHA todos os anos no país (DEWEY-MATTIA et al., 2018).
O norovírus é o agente etiológico responsável pelo maior número de casos de DTHA nos EUA, apesar de apresentar baixa letalidade, no período de 2009 a 2015, foram registrados 1.130 surtos, 27.623 doentes e sete mortes. A Salmonella spp, é o segundo agente etiológico mais incriminado em surtos de DTHA no país. No período de 2009 a 2015, foram notificados 896 surtos, com 23.662 doentes, e de três a quatro mortes todos os anos (DEWEY-MATTIA et al., 2018).
Nos EUA no período de 2009 a 2015, foram notificados 191 surtos de DTHA relacionados à contaminação por E.coli. Dentre estes, 2.387 indivíduos ficaram doentes, ocorreram 672 hospitalizações e 12 mortes (DEWEY-MATTIA et al., 2018).
O número de surtos causados por Listeria monocytogenes no período de 2009 a 2015 nos EUA foi baixo quando comparado com os outros patógenos envolvidos em surtos de DTHA no mesmo período. Entretanto, a taxa de mortalidade foi mais alta, com 74 mortes (DEWEY-MATTIA et al., 2018).
C. jejuni casou 140 surtos, 2.095 doentes e uma morte no período 2009 a 2015 nos EUA. Clostridium perfringens causou 108 surtos, 5.510 doentes, 16 hospitalizações e quatro mortes, nesse mesmo período. O vírus da hepatite A foi o segundo agente viral com maior número de surtos de DTHA nos EUA, foram notificados 15 surtos, 260 indivíduos doentes e 170 hospitalizações. Dos agentes parasitários, o Cryptosporidium spp foi o parasita que
mais casou surtos no país, foram notificados 10 surtos, com 160 doentes e seis hospitalizações (DEWEY-MATTIA et al., 2018).
União Europeia (UE)
O norovírus é uma das principais causas de doenças alimentares na UE, chegando a 15 milhões de casos todos os anos, juntamente com Campylobacter jejuni com mais de 5.000 milhões de casos. Relatórios do Centro Europeu de Prevenção e Controle de Doenças (ECDC) da UE, apontam que Campylobacter jejuni e Salmonella spp são os patógenos bacterianos que mais causam surtos de DTHA na União Europeia. No período de 2012 a 2020, os casos de C. jejuni pelo consumo de carne de frango permaneceu alto entre os países europeus. Somente em 2020, percebe-se uma redução do número de casos. Os casos de Salmonella spp também aumentaram no período 2014 a 2016 na UE, em 2014 foram 88.715, em 2018 são 91.857 casos registrados e 2020 os números reduziram para 52.702 (ECDC, 2014; ECDC, 2016; ECDC, 2018; ECDC, 2020).
A E. coli produtora da toxina shiga (STEC) e a Yersinia enterocolitica foram a terceira causa mais frequente de infecções bacterianas de origem alimentar na UE. No período de 2014 a 2020, os casos de DTHA por E. coli e Y. enterocolitica se mantiveram estáveis na Europa (ECDC, 2014; ECDC, 2016; ECDC, 2018; ECDC, 2020).
A Listeria monocytogenes é responsável pelos casos mais graves de DTHA na UE, listeriose é a doença de origem alimentar associada ao maior número de mortes, no período de 2014 a 2020, foram relatadas 853 mortes pela bactéria, seguido de Salmonella spp com cerca de 334 mortes (ECDC, 2014; ECDC, 2016; ECDC, 2018; ECDC, 2020).
Com relação as doenças parasitárias, Trichinella spiralis é o agente parasitário com maior número de notificações no período verificado, com um total de 603 indivíduos doentes, 205 hospitalizações e duas mortes em 2014 na UE (ECDC, 2014; ECDC, 2016; ECDC, 2018; ECDC, 2020)
Japão
No Japão, no período de 2014 a 2018 foram notificados 1.132 surtos de DTHA com
19.214 doentes. A maioria dos surtos de DTHA no Japão (40%) são originados por alimentos contaminados por bactérias. Os patógenos Salmonella spp, Campylobacter spp, Staphylococcus aureus, Clostridium perfringens, E. coli, Clostridium botulinum e Vibrio parahemolyticus são os principais microrganismos envolvidos em casos de DTHA no Japão. Um estudo feito no período de 2000 a 2018 mostra que o número de casos de DTHA com o envolvimento de Salmonella spp, S. aureus e E. coli diminuiu no país. O norovírus e Campylobacter spp foram os agentes com o maior número de casos de DTHA no Japão no período de 2000 a 2018. Clostridium perfringens, diferente dos outros países apresenta um número relativamente alto de casos no Japão (LEE et al., 2021).
Coréia
Diferente dos outros países, na Coréia a Escherichia coli no período de 2016 a 2022,
foi o patógeno responsável pelo maior número de surtos de DTHA com um total de 6.265 casos. O norovírus no mesmo período foi o patógeno viral mais envolvido em surtos de origem alimentar no país, com 3.573 notificações. Em 2018, os casos de DTHA relacionados com Salmonella spp aumentaram 10 vezes quando comparado aos casos notificados em 2016. Staphylococcus aureus, embora apresente um aumento no número de casos, em 2016 foram notificados quatro surtos e em 2022 foram registrados 111 casos, ainda assim, é o
patógeno bacteriano com menor número de casos de DTHA na Coréia. Todos os dados podem ser observados na Tabela 3 (COREA, 2022).
Brasil
No Brasil, a vigilância de surtos de DTHA teve início em 1999. Os surtos de DTHA
constituem eventos de saúde pública que representa uma ameaça á saúde da população. A Resolução RDC de n°724 de 01 de julho de 2022 dispõe sobre os padrões microbiológicos para os alimentos e suas aplicações e a Instrução Normativa IN de n°161 de 01 de julho de 2022 estabelece os padrões microbiológicos para os alimentos (BRASIL, 2022).
No período de 2012 a 2021 foram notificados no Brasil, 6.347 surtos de DTHA com 610.684 indivíduos expostos, 104.843 doentes, 13.446 hospitalizações e 89 mortes. Destes, somente em 1.559 foi possível realizar a identificação do agente etiológico envolvido. Escherichia coli foi responsável pelo maior número de surtos correspondendo a 29,6% dos casos, seguido dos patógenos Staphylococcus aureus 12,9%, Salmonella spp 11,2% e Bacillus cereus 7,2%. Os demais agentes etiológicos registrados nesse período foram norovírus 5,2%, rotavírus 4,4%, Clostridium perfringens 3,0%, E. coli Enteropatogênica (EPEC) 2,8%, T. cruzi 2,0%, Shigella sonnei 1,7%, vírus da hepatite 1,5% e outros 13,7%
(BRASIL, 2022).
Diferente do Brasil e da Coréia em que a E. coli é a principal causa de surtos de DTHA, nos Estados Unidos, o norovírus lidera o número de casos de DTHA. No período de 2009 a 2015 foram 1.130 surtos, 27.623 doentes e sete mortes pelo vírus nos EUA. O mesmo cenário pode ser observado na União Europeia, o número de casos de DTHA por norovírus chega a 15 milhões todos os anos. No Japão o norovírus é o agente viral associado ao maior número de casos de DTHA. O número de surtos por norovírus no Brasil, de 2012 a 2021 corresponde a 89 (5,2%) de casos. Na Coréia o norovírus é a segunda maior causa de surtos de DTHA, de 2016 a 2022 foram notificados 3.705 casos (BRASIL, 2022; CORÉIA, 2022; DEWEY-MATTIA et al., 2018; ECDC, 2015; LEE et al., 2021).
O número de mortes relatadas no Brasil causadas por DTHA no período de 2012 a 2021 consistiu em 89 mortes. A maioria dos óbitos, 15, foram causados por E.coli, seguido de Salmonella spp com quatro mortes e Salmonella Enteritidis com uma morte, S. aureus três mortes, Shigella flexneri uma morte e uma morte relatada por Shigella spp. Listeria monocytogenes causou quatro mortes em 2021 e Clostridium botulinum três mortes. Em 2018 o consumo de água contaminada acarretou um surto de hepatite A com sete indivíduos doentes e sete mortes na região Sudeste do Brasil. Uma morte foi relatada em 2019 por Trypanosoma cruzi. Entretanto, o agente etiológico na maioria das notificações de óbitos no período de 2012 a 2021 no Brasil não foram identificados, dos 89 óbitos relatados, em 49 o agente etiológico não foi identificado (BRASIL, 2022).
Apesar da comprovada relação das várias doenças pela ingestão de alimentos e água contaminados no Brasil com número de surtos e doentes apresentados, pouco se conhece a real dimensão do problema, devido à falta de dados. O número de notificações de surtos de DTHA no Brasil é muito menor quando comparado a outras partes do mundo como União Europeia e Estados Unidos, porém, não significa que o número de casos seja expressivamente maior nesses países, mas que possivelmente a vigilância epidemiológica seja mais estruturada e que essa diferença seja referente as subnotificações da doença (BRASIL, 2022).
A investigação de surtos de DTHA no Brasil ocorre com base na confirmação dos critérios clínico epidemiológico para o esclarecimento do caso, no entanto, a partir de 2012 nota-se uma pequena redução desse critério na conclusão de investigações de surto de DTHA (28%) e, consequentemente o aumento de casos inconclusivos (40%). O critério laboratorial
clínico bromatológico ideal para identificação dos agentes etiológicos envolvidos nos surtos, se manteve estável nesse período, porém baixo (5%), sendo possível observar que a maior parte dos surtos o agente etiológico não foi possível ser identificado (BRASIL, 2022).
Ainda que a investigação dos surtos de DTHA pelo o critério clínico epidemiológico permita uma alta identificação do número de casos, o critério laboratorial é extremamente importante para o encerramento dos surtos, porque possibilita traçar um perfil epidemiológico dos agentes etiológicos circulantes, com maior prevalência em surtos de DTHA, possibilitando melhores ações de diagnostico, prevenção e controle (BRASIL, 2022).
CONCLUSÃO
Com base nas informações obtidas nesse estudo, conclui-se que os principais agentes etiológicos envolvidos em surtos de DTHA em diferentes regiões do mundo são norovírus, Salmonella spp, E. coli, Campylobacter spp, Staphylococcus aureus e Listeria monocytogenes.
Os principais patógenos envolvidos em surtos de DTHA em países desenvolvidos são norovírus, Campylobacter jejuni, e Salmonella spp. Com relação a países em desenvolvimento, a E. coli e Salmonella spp são as causas mais frequentes de surtos. A Salmonella spp e E. coli são uma preocupação de saúde pública em países de alta e baixa renda. Os patógenos responsáveis pelo o maior número de mortes de DTHA no Brasil no período de 2012 a 2021 são E. coli e Salmonela spp.
Considerando o índice de casos de DTHA em diferentes regiões do mundo, e o impacto que essas doenças podem causar principalmente em países em desenvolvimento, com maior situação de vulnerabilidade, destaca-se a importância dos estudos sobre as DTHA para ações e medidas de controle sejam adotadas por Autoridades Sanitárias.
REFERÊNCIAS
ANTUNES, P.; NOVAIS, C.; PEIXE, L. Food-to-Humans Bacterial Transmission.
Microbiology Spectrum, v. 8, n. 1, p. 8.1.1, 16 jan. 2020.
BRASIL. Ministério da Saúde. Doenças de Transmissão Hídrica e Alimentar. DTHA. 2022. Disponível em: . Acesso em: 28 fev. 2023.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. ANVISA. Resolução – RDC N° 724, de 1° de julho de 2022. Dispõe sobre os padrões microbiológicos e suas aplicações. Diário Oficial da União, Brasília, DF, 06 jul. 2022. Disponível em < https://www.in.gov.br/en/web/dou/-/resolucao-rdc-n-724-de-1-de-julho- de-2022-413364812 > Acesso em: 20 nov. 2022.
BRASIL. Ministério da Saúde. Agência Nacional de Vigilância Sanitária. ANVISA. Instrução Normativa – IN N° 161, de 1° de julho de 2022. Estabelece os padrões microbiológicos dos alimentos. Diário Oficial da União, Brasília, DF, 06 jul.2022. Disponível em > https://in.gov.br/en/web/dou/-/instrucao-normativa-in-n-161-de-1-de- julho-de-2022-413366880 >Acessado em: 20 nov. 2022
BRASIL. Ministério da Saúde. Surtos de Doenças de Transmissão Hídrica e Alimentar informe 2022. Ministério da Saúde Secretaria de Vigilância em Saúde Departamento de Imunização e Doenças Transmissíveis Coordenação-Geral de Vigilância de Zoonoses e Doenças de Transmissão Vetorial. 2022. Disponível em: Acesso em: 26 nov. 2022.
BRASIL. Ministério da Saúde. Banco de Dados de Surtos de DTHA-2000 a 2021. 2022. Disponível em: https://www.gov.br/saude/pt-br/assuntos/saude-de-a-a-z/d/dtha/dtha.
Acesso em: 12 out. 2022.
CORÉIA. Estatísticas de intoxicação alimentar: país de segurança alimentar. Disponívelem:. Acesso em: 29. 2022.
DEWEY-MATTIA, Daniel; MANIKONDA, Karunya; HALL, Aron J.; et al. Surveillance for Foodborne Disease Outbreaks — United States, 2009–2015. MMWR Surveillance Summaries, v. 67, n. 10, p. 1–11, 2018.
ECDC. European Centre for Disease Prevention and Control. European Food Safety Authority. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food‐borne outbreaks in 2014.EFSA Journal, v. 13, n. 12, p. 4329, 2015. Disponível em: Acesso em: 20 nov. 2022.
ECDC. European Centre For Disease Prevention and Control. The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA Journal, v. 15, n. 12, p. e05077, 2017. Disponível em: https://efsa.onlinelibrary.wiley.com/doi/abs/10.2903/j.efsa.2017.5077 > Acesso em: 20 nov. 2022.
ECDC. European Centre for Disease Prevention and Control. The European Union One Health 2018 Zoonoses Report. Disponível em:
. Acesso em: 7 dez. 2022.
ECDC. European Centre For Disease Prevention and Control. European Food Safety Authority. The European Union One Health 2020 Zoonoses Report. EFSA Journal, v. 19,
n. 12, 2021. Disponível em: https://data.europa.eu/doi/10.2903/j.efsa.2021.6971. Acesso em: 7 dez. 2022.
LEE, H.; YOON, Y. Etiological Agents Implicated in Foodborne Illness World Wide.
Food Science of Animal Resources, v. 41, n. 1, p. 1–7, jan. 2021.
NYACHUBA, D. G. Foodborne illness: is it on the rise?: Nutrition Reviews, Vol. 68, No. 5. Nutrition Reviews, v. 68, n. 5, p. 257–269, 28 abr. 2010.
SWITAJ, T. L.; WINTER, K. J.; CHRISTENSEN, S. R. Diagnosis and Management of Foodborne Illness. American Family Physician, v. 92, n. 5, p. 358–365, 1 set. 2015.
WHO. World Health Organization. Estimates of the global burden of foodborne diseases: foodborne disease burden epidemiology reference group 2007-2015. Geneva, p 72. 2015. Disponível em: https://apps.who.int/iris/handle/10665/199350. Acesso em: 16 out. 2022.
Capítulo de livro publicado no Livro da IV Mostra dos Trabalhos de Conclusão de Curso da Especialização em Vigilância Laboratorial em Saúde Pública. Para acessa-lo clique aqui.
doi.org/10.53934/9786599965821-03
Este trabalho foi escrito por:
Wellington Rudnei Athanazio1, Marina von Atzingen2
1Estudante do Curso de Especialização Vigilância Laboratorial em Saúde Pública –SUS/SP – IAL; E-mail: [email protected]
2Docente/Pesquisadora Científica do Núcleo de Ciências Biomédicas, Centro de Laboratório Regional de Ribeirão Preto VI – Instituto Adolfo Lutz (IAL-CLR-VI-RP).
RESUMO
Mutações são acidentes ou erros que acontecem ocasionalmente em um organismo que possui material genético, sejam eles DNA ou RNA. Acompanhar essas variações nos vírus tem tornado possível ao ser humano entender sua disseminação, transmissibilidade e virulência. Em 2019, emergiu um coronavírus da família Coronoviridae, o SARS-CoV-2, causando nos seres humanos infecções abruptas, doença essa chamada de COVID-19. Ele sofreu diversas mutações ao longo do período pandêmico, mas elas não pararam. Suas mutações formaram variantes virais capazes de mudar até mesmo a sintomatologia da doença. Este trabalho tem como objetivo, realizar uma revisão bibliográfica, com o auxílio de bancos de dados governamentais mundiais e repositórios como Scielo, Pubmed, Organização Mundial da Saúde, Ministério da Saúde e demais espalhados pelos continentes, a fim de explorar sua origem, mutações, linhagens, variantes e cepas, deste vírus que circula desde novembro de 2019 entre os seres humanos; sua patogenicidade e características fenotípicas. Com isso, compreender as perspectivas atuais e entender como se desenvolverá este vírus ao longo do tempo, em um período pós-vacinal, tem sido um desafio para os pesquisadores que trabalham arduamente para compreender como frear a infecção que este vírus provoca e tentar definir se uma cepa única circulará sobre a população do mundo, ou se novas variantes mais poderosas poderão surgir ao longo dos anos.
Palavras–chave: Epidemias; Infecções por coronavírus; SARS-CoV-2; Vigilância;; Vírus RNA
INTRODUÇÃO
O novo coronavírus, SARS-CoV-2, foi responsável pela morte de mais de seis milhões de pessoas em todo mundo e até o presente momento observa-se casos de óbito. Em menos de três meses de seu aparecimento na China, a COVID-19 tornou-se pandêmica causando mortes, crise econômica e o colapso de sistema de saúde de vários países (MENEZES, 2020).
Depois de sua emergência em Wuhan, outras cidades foram identificadas como epicentro da doença. À medida que o vírus se espalhava pelos continentes, cidades de outros países, se destacavam como epicentro pandêmico, como foi o caso da Itália, França, Espanha,
Inglaterra, EUA. Para se ter ideia da velocidade da contaminação e da gravidade deste patógeno, em 31 de março de 2020 havia 760.040 casos confirmados e 40.842 mortos. Em um período de aproximadamente 6 meses, após estes casos, houve um salto para 32.925.668 de casos confirmados e 995.352 mortes (SOUZA et al., 2021).
Este vírus pertence a uma classe de vírus zoonóticos, compostos por RNA que acometem uma variedade de animais, incluindo aves e mamíferos. Destes, apenas sete são reconhecidos como patogênicos para humanos. Nos últimos 20 anos, três deles foram responsáveis por grandes epidemias virulentas de Síndrome Respiratória Aguda Grave (SRAG), sendo elas: a epidemia de SARS de Hong Kong (China) em 2003, a síndrome respiratória do Oriente Médio (MERS), que emergiu na Arábia Saudita em 2012 e atualmente o SARS-CoV-2 (da sigla em inglês coronavírus 2 da síndrome respiratória aguda grave) que em 31 de dezembro de 2019 emergiu em Wuhan, na China (CORMAN et al., 2018; LI et al., 2020).
Explorar essas variantes, como elas surgem e de que maneira se desenvolvem é o propósito desta revisão bibliográfica, abordando suas variantes de preocupação, de interesse e de alta consequência, pois estudos como este podem auxiliar no monitoramento de possíveis futuras epidemias, bem como presumir o surgimento do que chamam de “novas onda” (quando novos surtos de determinada doença ressurge), na produção de uma vacina eficaz e até mesmo compreender quais estruturas virais podem estar mais relacionadas às suas mutações. Descrever as mutações, linhagens, variantes e cepas diferentes deste vírus que circula há quase três anos no mundo, desde seu primeiro caso de transmissão na cidade de Wuhan, na China; entender as origens e rearranjos moleculares que o SARS-Cov-2 tem sofrido ao longo do tempo, para assim, compreender as perspectivas atuais e observar como este vírus se desenvolverá ao longo do tempo, de um período pré ao pós-vacinal.
MATERIAL E MÉTODOS
A elaboração deste trabalho teve como premissa, realizar uma revisão bibliográfica, com o auxílio de bancos de dados governamentais mundiais e repositórios como Scielo, Pubmed, Organização Mundial da Saúde, Ministério da Saúde e demais espalhados pelos continentes. Os artigos foram selecionados e reunidos a partir das palavras-chave: “SARSCoV-2″, “Infecções por coronavírus”, “Vigilância”, “Epidemias”, “Vírus RNA”, a fim de facilitar a busca por artigos que contribuam para elaboração deste trabalho.
RESULTADOS E DISCUSSÃO
O SARS-CoV-2, ou novo coronavírus, é um vírus da família Coronoviridae, envelopado, aproximadamente esférico, que possui genoma de RNA positivo não segmentado de fita simples, codificando quatro proteínas principais: glicoproteína S (Spike), proteína do envelope (E), glicoproteína da membrana (M), e proteína nucleocapsídeo (N) (KHALIL & KHALIL, 2020). Foi isolado pela primeira vez em 1937, mas somente em 1965 foi nomeado de corona quando sua morfologia, em aspecto de coroa, foi observada a partir de uma análise microscópica das proteínas de sua superfície (LIMA, 2020; STEPHENS et al, 2010). Possui menos de 30.000 nucleotídeos em seu genoma, formados por moléculas de ribose, um ácido fosfórico e uma base nitrogenada. A identificação de cerca de 29 proteínas
virais diferentes facilitou a compreensão das estruturas que compõem suas características (UZUNIAN, 2020)
A transmissão se dá a partir da inalação de gotículas e/ou aerossóis, de uma pessoa contaminada, projetados em forma de espirros, tosses ou em procedimentos hospitalares. Depois de inalada, a partícula viral do SARS-CoV-2 se conecta com receptor da célula humana através da proteína Spike que possui afinidade com um receptor protéico humano, chamado ACE2 (Enzima Conversora de Angiotensina 2). Os sintomas clínicos da infecção pelo novo coronavírus também pode variar desde uma Síndrome Gripal (SG), com sinais e sintomas considerados leves como tosse, congestão nasal e fadiga, até uma SRAG, com sintomas adicionais como, saturação de O2 ≤ 93%, frequência respiratória ≥ 30 rpm, pressão arterial de oxigênio (PaO2) /fração inspirada de oxigênio (FIO2) < 300, linfopenia e edema alveolar; 80% de casos leves e assintomáticos. Aproximadamente 49% dos casos críticos desenvolvem insuficiência respiratória, choque séptico chegando a óbito (REINHARDT, 2022; LI et al., 2020; UZUNIAN, 2020; VALVERDE, 2021; BAGGIO et al., 2021; ISER et al., 2020).
À medida que os vírus infectam um indivíduo, circulam livremente em uma
população, podem sofrer modificações em suas estruturas durante a multiplicação, gerando novas cepas, linhagens ou ainda variantes virais. As mutações são características que definem a taxa de sobrevivência de vírus compostos por RNA, acumulando-as por cada ciclo de cópia, podendo ocorrer em horas, criando em um único hospedeiro, vários vírus diferentes (GRUBAUGH et al, 2020). Isto pode ser vantajoso do ponto de vista de a variante modificada, originar uma linhagem viral com pouca força ou capacidade de virulência aumentando sua disseminação, ou seja, quanto menos virulento mais transmissível é o vírus, porque é mais fácil transmitir um vírus através de indivíduos assintomáticos, que se sentem bem clinicamente (SOARES, 2020).
Assim sendo, quando uma variante obtém sucesso em suas mutações, ao se difundir para outros países ou continentes, se espalhando para uma grande quantidade de pessoas ela se torna uma linhagem. Todavia, uma linhagem é uma variante, mas nem toda variante se torna uma linhagem, isso porque mesmo que haja mutações, se a linhagem não for capaz de se propagar com eficiência ela desaparece.Variantes muito semelhantes às linhagens de onde se ramificaram o que acaba sendo expresso em sua nomenclatura, são classificadas sublinhagens, sendo acrescentado “.1” ou “.2” para indicar que é da mesma estirpe. Como é o caso das sublinhagens da variante ômicron que recebem o nome BA.1 => BA.1.1; BA.1.2 e assim por diante (BUTANTAN, 2022a).
Já quando é alterado ao menos uma das características observáveis ou fenotípicas de um vírus, são denominadas cepas. Para uma melhor compreensão, é quando uma porção viral desenvolve uma capacidade de transmissão, resposta imunológica ou produz sintomas em um indivíduo que difere dos observados concebidos por seu ascendente (VALVERDE, 2021). Para amparar a compreensão das mutações, as variantes foram classificadas conforme sua importância facilitando a organização das ações no combate às pandemias. A Organização Mundial da Saúde e equipes de especialistas do mundo todo, monitoram as mudanças que ocorrem no vírus, para que se mutações significativas forem identificadas, os países possam ser notificados para adotarem medidas de contenção, para evitarem a propagação de uma nova linhagem (OPAS, 2021).
Conforme descrito por Varella (2022), variantes de interesse têm por definição os vírus que, ao sofrer mutação, desenvolvem uma capacidade maior de transmissão. Ou seja,
segundo a OPAS (2021), é considerada uma variante de interesse, se ao ser comparada à sua variante original, o seu genoma tiver informações capazes de modificar o fenótipo do vírus, se tiver sido causadora de transmissão comunitária, se de outra forma, for avaliada pela OMS como tal, em consonância com grupos de pesquisa sobre a evolução do SARSCoV-2.
Há também as variantes de preocupação. As que foram mais destacadas no mundo todo foram: Alfa, Beta, Gama, Delta e Ômicron. Existe a preocupação de que mutações relacionadas a essas variantes possam provocar um escape do sistema imunológico, doenças mais graves e aumento na transmissibilidade; por isso a importância em se classificar e estudar estas variantes (FOX, 2021a). E por fim, as variantes de alta consequência são as que respondem menos aos imunizantes, causam infecções mais graves e hospitalizações. Até o momento não foram identificadas nenhuma deste gênero (BUTANTAN, 2021b).
A OPAS criou uma rede entre laboratórios de vários países com capacidade de sequenciamento interno e externo com treinamentos e ações de suporte para geração de sequenciamento genômico do SARS-CoV-2. O compartilhamento destas informações na plataforma GISAID (do inglês, Global Initiativeon Sharing All Influenza Data) também é incentivado (OPAS, 2020).
GISAID é uma plataforma cientifica iniciada em 2008 após eventos da pandemia da gripe aviaria (H5N1) de 2006. A plataforma realiza o compartilhamento de dados entre os Centros Colaboradores da OMS e os Centros Nacionais de Influenza disponibilizando atualizações e recomendações semestrais do vírus da vacina contra a gripe pelo Sistema Global de Vigilância e Resposta à Gripe da OMS (GISRS) e de patógenos correlacionados. Este compartilhamento viabilizou a troca de informações entre os órgãos governamentais e a GISAID por onde foi possível, por exemplo, o compartilhamento de 125 mil sequências genômicas virais do novo coronavírus (FERREIRA, 2021).
CONCLUSÕES
A capacidade de transmissão do novo coronavírus é inquestionável. Sua virulência e letalidade variam muito dependendo do indivíduo que acomete, entretanto cerca de 20% da população pode desenvolver algumas das formas moderadas, graves ou críticas. Em 2019, o uso de máscaras, o isolamento social e as mudanças na questão de higienização das mãos, tiveram como intuito evitar a disseminação descontrolada do vírus, o que poderia facilitar suas mutações e causar um colapso global nos sistemas de saúde, como foi o caso de alguns países.
O fato de a maioria das mutações acontecerem na estrutura da proteína spike facilitou a criação de uma vacina eficaz no combate a infecção, através de métodos já conhecidos para confecção de vacinas. Com a chegada da vacina em 2021, algumas medidas profiláticas foram sendo desobrigadas. Pensava-se que a vacina poderia impedir, de maneira generalizada a disseminação do vírus. No entanto, mesmo com a facilidade da vacina atualmente, ainda pouco mais de 51% da população brasileira tomaram suas doses reforço para novo coronavírus. Isto pode favorecer o aparecimento de novas variantes, capazes de fugirem da resposta imunológica de um indivíduo vacinado. Uma pessoa não vacinada pode ser um anfitrião caloroso para este tipo de vírus.
A facilidade de serem compiladas cópias mutantes, dentro de uma pessoa, é imensurável e isso pode possibilitar o aparecimento de linhagens mais agressivas. A variante ômicron demonstrou que veio pra ficar, pois a maioria de suas linhagens tende a sofrer
mutações com muita facilidade. Sempre haverá uma forma de os vírus evoluírem, e o SARS- CoV-2 não é tão diferente. Pode ser protelada, mas nunca contida, podendo ou não ser irrelevante.
A ideia de convergência evolutiva deixa claro que existe uma tendência nas mutações de elas surgirem e serem parecidas, mesmo em locais distintos do globo terrestre, sem nunca terem estado próximas. Com isso, é fundamental manter a vigilância genômica, pois pode favorecer a identificação das mutações de forma bem mais rápida e até facilitar a produção de vacinas adaptadas a fim de alcançarem uma imunização mais completa da população.
REFERÊNCIAS
BAGGIO, J. A. O. et al.; Síndrome Respiratória Aguda Grave (SRAG) Causada por
COVID-19: um fator regional. Arquivos Brasileiros de Cardiologia, Arapiraca, v. 117, n. 5, p. 976-977, 22 nov. 2021. Sociedade Brasileira de Cardiologia. http://dx.doi.org/10.36660/abc.20210803.
BUTANTAN, Instituto. A velocidade com que foi criada a vacina da Covid-19 é motivo de preocupação? Especialista do Butantan responde. 2021a. Portal do Butantan. Disponível em: https://butantan.gov.br/covid/butantan-tira-duvida/tira- duvida-noticias/a-velocidade-com- quefoi-criada-a-vacina-da-covid-19-e-motivo-de-preocupacao-especialista-do-butantan- responde. Acesso em: 30 nov. 2022.
BUTANTAN, Instituto. Como surgem as variantes da Covid-19: entenda a diferença entre linhagens, sublinhagens e recombinantes. 2022a. Portal do Butantan. Disponível em: https://butantan.gov.br/covid/butantan-tira-duvida/tiraduvida-noticias/como-surgem- asvariantes-da-covid-19-entenda-a-diferenca-entrelinhagens-sublinhagens-e-recombinantes. Acesso em: 18 nov. 2022
BUTANTAN, Instituto. Por que acontecem mutações do SARS-CoV-2 e quais as diferenças entre cada uma das variantes. 2021b. Portal do Butantan. Disponível em: https://butantan.gov.br/noticias/por-que-acontecem-mutacoes-do-sars-cov-2-equais- asdiferencas-entre-cada-uma-das-variantes. Acesso em: 22 nov. 2022.
BUTANTAN, Instituto. Vacinação está impedindo avanço das variantes da Covid-19 no Brasil, explicam cientistas do Butantan. 2022b. Portal do Butantan. Disponível em: https://butantan.gov.br/noticias/vacinacao-esta-impedindo-avanco-das-variantes-da-covid- 19no-brasil-explicam-cientistas-do-butantan. Acesso em: 23 nov. 2022.
CORMAN, V. M. et al. Hosts and Sources of Endemic Human Coronaviruses. Advances in Virus Research, New York, v. 100, p. 163-188, 2018. DOI: 10.1016/ bs.aivir.2018.01.001.
Disponível em: https://www.sciencedirect.com/science/article/pii/S0065352718300010?via%3Dihub. Acesso em: 16 nov 2022.
FERREIRA, V. Fiocruz integra plataforma de dados genômicos sobre influenza e novo coronavírus. 2021. Fundação Osvaldo Cruz – Fiocruz. Disponível em:
https://portal.fiocruz.br/noticia/fiocruz-integra-plataforma-de-dados-genomicos- sobreinfluenza-e-novo-coronavirus. Acesso em: 01 dez. 2022.
FOX, Maggie. Ómicron, mu, delta, lambda y otras: una por una, todas las variantes del coronavirus identificadas hasta el momento. 2021a. Cable News Network – CNN Espanha. Disponível em: https://cnnespanol.cnn.com/2021/11/29/variantes-coronavirus- omicronidentificadas-trax/#0. Acesso em: 22 nov. 2022.
GRUBAUGH, Nathan D.; PETRONE, Mary E.; HOLMES, Edward C. We shouldn’tworry when a vírus mutates during disease out breaks. Nature, [S. l.], ano 2020, n. 5, p. 529-530, 18 fev. 2020. DOI https://doi.org/10.1038/s41564-020-0690-4.
ISER, B. P. M.; et al. Definição de caso suspeito da COVID-19: uma revisão narrativa dos sinais e sintomas mais frequentes entre os casos confirmados. Epidemiologia e Serviços de Saúde, Tubarão, v. 29, n. 3, p. 1-11, abr. 2020. FapUNIFESP (SciELO). http://dx.doi.org/10.5123/s1679-
49742020000300018.
KHALIL, O. A. K.; KHALIL, S. da S. SARS-CoV-2: taxonomia, origem e constituição. Revista de Medicina, [S. l.], v. 99, n. 5, p. 473-479, 2020. DOI:
10.11606/issn.1679-9836.v99i5p473-479. Disponível em: https://www.revistas.usp.br/revistadc/article/view/169595. Acesso em: 4 nov. 2022.
LI, X. et al. Molecular immune pathogenesis and diagnosis of COVID-19. Journal Of Pharmaceutical Analysis, [S.L.], v. 10, n. 2, p. 102-108, abr. 2020. Elsevier BV. http://dx.doi.org/10.1016/j.jpha.2020.03.001.
LIMA, C. M. A. de O. Informationaboutthe new coronavirusdisease (COVID-19). Radiologia Brasileira, Rio de Janeiro, v. 53, n. 2, p. 5-6, abr. 2020. FapUNIFESP (SciELO). http://dx.doi.org/10.1590/0100-3984.2020.53.2e1.
MENESES, A. S. de. COVID-19 NATURAL HISTORY AND ITS THERAPEUTIC
RELATIONSHIPS. SciELO Preprints, 2020. DOI: 10.1590/SciELOPreprints.733. Disponível em: https://preprints.scielo.org/index.php/scielo/preprint/view/733. Acesso em: 1 nov. 2022.
OPAS, Organização Pan Americana de Saude -. OMS anuncia nomenclaturas simples e fáceis de pronunciar para variantes de interesse e de preocupação do SARS-CoV-2. 2021. OPAS. Disponível em: https://www.paho.org/pt/noticias/1-6- 2021-oms-anuncia-nomenclaturas-simples-efaceis- pronunciar-para-variantesinteresse-e. Acesso em: 18 nov. 2022.
OPAS, Organização Pan Americana de Saude -. Rede Regional de Vigilância Genômica de COVID-
- Disponível em: https://www.paho.org/pt/node/4951/rede-regional-vigilanciagenomica- covid-19. Acesso em: 23 nov. 2022.
REINHARDT, É. L. Transmissão da COVID-19: um breve reexame das vias de transmissão por gotículas e aerossóis. Revista Brasileira de Saúde Ocupacional, São Paulo, v. 47, p. 1-9, 06 abr. 2022. FapUNIFESP (SciELO). http://dx.doi.org/10.1590/2317-6369000000221.
SOARES, F. COVID-19: É possível matar o que não está vivo? Ciencia Viva, 26/07/2020. Disponível em: https://cidadeverde.com/cienciaviva/109838/covid-19-epossivel-matar-o-que-naoesta-vivo. Acesso em: 29 out. 2022.
SOUZA, A. S. R. et al. General aspects of the COVID-19 pandemic. Revista Brasileira de Saúde Materno Infantil, Recife, v. 21, n. 1, p. 29-45, fev. 2021. FapUNIFESP (SciELO). http://dx.doi.org/10.1590/1806-9304202100s100003.
STEPHENS, P. R. S. et al. In: FUNDAÇÃO OSWALDO CRUZ. Conceitos e Metodos para Formação de Profissionais em Laboratorios de Saúde. Rio de Janeiro: [s. n.], 2010. v. 4, cap. Virologia, p. 185- 186.
UZUNIAN, Armênio. Coronavirus SARS-CoV-2 and Covid-19. Jornal Brasileiro de Patologia e Medicina Laboratorial, São Paulo, p. 1-4, 25 set. 2020. GN1 Genesis Network. http://dx.doi.org/10.5935/1676-2444.20200053.
VALVERDE, R. O que são mutações, linhagens, cepas e variantes? Agência Fiocruz de
Notícias, Rio de Janeiro, 29, jun. 2021. Disponível em: 36 https://portal.fiocruz.br/noticia/o-quesao- mutacoes-linhagens-cepas-e-variantes-1. Acesso em: 01 out 2022.
VARELLA, Drauzio. COVID-19: CONHEÇA OS PRINCIPAIS SINTOMAS DE CADA
VARIANTE. 2022. Portal Drauzio Varella – Informação sobre saúde para todos. Disponível em: https://drauziovarella.uol.com.br/coronavirus/covid-19-conheca-osprincipais-sintomas-de- cadavariante/. Acesso em: 22 nov. 2022.
Capítulo de livro publicado no Livro da IV Mostra dos Trabalhos de Conclusão de Curso da Especialização em Vigilância Laboratorial em Saúde Pública. Para acessa-lo clique aqui.
doi.org/10.53934/9786599965821-02
Este trabalho foi escrito por:
Gustavo Carvalho Amorim1; Carlos Roberto Prudêncio2
1Estudante do Curso de Especialização em Vigilância Laboratorial em Saúde Pública – Centro de Imunologia – Instituto Adolfo Lutz; E-mail: [email protected] 2Docente/pesquisador do Centro de Imunologia – Instituto Adolfo Lutz.
Resumo: A pandemia da COVID-19 trouxe inúmeros desafios para a sua mitigação ao redor do globo. A tecnologia de imunoprecipitação de fagos seguida pelo sequenciamento de alto rendimento (PhIP-Seq) propõe analisar o histórico imunológico individual e estudar interações antígeno-anticorpo por meio de fagos que expressam proteínas e peptídeos de interesse. Esse trabalho tem por objetivo otimizar e padronizar a técnica de titulação e expansão de uma biblioteca de antígenos de coronavírus expressos em fagos T7 (CoronaScan), bem como a quantificação de imunoglobulinas IgG (IgG) de amostras em soro humano por ELISA, que, combinadas, permitirão o aprimoramento de análises e estudos imunológicos e epidemiológicos. A cepa E. coli BLT5403 foi usada como hospedeira nos ensaios de titulação e expansão da biblioteca CoronaScan em placas ágar LB. Para a quantificação de IgG, uma curva padrão foi construída a partir de uma solução de IgG purificada e com concentração conhecida, juntamente a determinação de IgG de uma amostra de soro, que deverá ser utilizado como controle nos futuros ensaios. Após a expansão, biblioteca CoronaScan (1,5×105 ± 0,7 pfu/mL) passou a ter a titulação de 1,5×1010 ± 2,12 pfu/mL, enquanto a concentração de IgG adequado para o método se encontrou na diluição de 1/1.000.000. Apesar da otimização e a da padronização de tais técnicas terem sido realizadas, novos ensaios de quantificação de IgG pelo método de ELISA serão necessários para assegurar a reprodutibilidade. Além disso, recomenda-se a realização do controle da qualidade da biblioteca CoronaScan a fim de analisar a sua diversidade de peptídeos.
Palavras–chave: coronavírus; PhIP-Seq; T7; vigilância epidemiológica
INTRODUÇÃO
A pandemia da COVID-19 (Coronavirus disease 2019), doença causada pelo vírus SARS-CoV-2 (do inglês severe acute respiratory syndrome coronavirus 2), requereu um grande esforço mundial para o controle de sua disseminação. Em dezembro de 2022, dados da Organização Mundial da Saúde (OMS) confirmaram um pouco mais de 630 milhões de casos e 6,6 milhões de mortes ocasionados pela doença, que foi identificada pela primeira vez na cidade de Wuhan, na China (Herstein et al., 2021; Peng et al., 2021; Rahman et al., 2021).
Mutações aleatórias nas proteínas estruturais spike e do nucleocapsídeo podem não somente dificultar o diagnóstico como também abalar as estratégias da vigilância epidemiológica já instauradas. A tecnologia de imunoprecipitação de fagos seguida pelo sequenciamento de alto rendimento (NGS) (PhIP-Seq, do inglês Phage
Immunoprecipitation Sequencing) é uma ferramenta poderosa capaz de investigar respostas do sistema imune provocadas por infecções virais, doenças imunes e infecciosas que podem comprometer o sistema imunológico. A técnica, ao ser combinada com a biblioteca de fagos T7 coronavírus VirScan (CoronaScan) – que representa os peptídeos dos coronavírus humanos endêmicos (HCoV-NL63, HCoV-229E, HCoV-OC43 e HCoVHKU1), SARS- CoV-1, SARS-CoV-2 e MERS (Middle East Respiratory Syndrome), além dos peptídeos de coronavírus encontrados em morcegos (BatCoV-Rp3, BatCov-HKU3 e BatCov-279) -, permite a identificação de peptídeos reconhecidos por imunoglobulinas IgG (IgG) por meio da imunoprecipitação mediada por esferas magnéticas sensibilizadas com proteínas A e G, com posterior amplificação das sequências codificantes por reação em cadeia da polimerase (PCR) e sequenciamento de nova geração (NGS) (Mohan et al., 2018; Morgenlander et al., 2021; Wiegand et al., 2022; Xu et al., 2015).
A biblioteca de fagos tem como base o phage display desenvolvido a partir de fagos T7, que apresenta vantagens frente ao fago M13, como maior estabilidade na presença de inserto, alta taxa de replicação em Escherichia coli, resistência a condições extremas e entre outras. (Deng et al., 2018; Krumpe & Mori, 2014; Tan et al., 2016; Yue et al., 2022).
Portanto, este trabalho tem como objetivo a otimização e padronização a titulação e expansão da biblioteca de fagos CoronaScan, e a padronização do ensaio de ELISA para a quantificação de IgG presentes no soro dos pacientes que serão utilizados nos experimentos de imunoprecipitação com a biblioteca.
MATERIAL E MÉTODOS
Os ensaios foram realizados de acordo com os protocolos descritos no Manual T7Select® System da Novagen (2011) e por Mohan e colaboradores (2018). Os resultados foram expressos como média e desvio-padrão (±) de pelo menos dois ensaios independentes. A titulação requereu E. coli BLT5403 com OD600 = 1,0 u.a. cultivada em meio
M9LB (NOVAGEN, 2011). As bactérias foram crescidas neste meio suplementado com x (concentração) ampicilina a 37° C sob agitação constante de 200 RPM. As células hospedeiras foram incubadas por 8 min com 100 µL da biblioteca CoronaScan previamente diluída em meio LB nas diluições de 102 a 109. Em seguida, 3 mL de meio top agarose (20 g de meio LB e 0,6 de agarose low melting em volume final de 1 L) foi adicionado para cada diluição, sendo essa mistura plaqueada sob uma camada de LB ágar com ampicilina a 50 µg/mL em uma superfície plana e incubada overnight a temperatura ambiente (TA). A titulação (pfu/mL) foi determinada por meio da contagem das unidades formadoras de placas (pfu) na placa LB contendo a maior diluição em que as placas de fagos puderam ser contadas de modo isolado. A estimativa da concentração da biblioteca foi auxiliada por meio da seguinte fórmula: Titulação (pfu/mL) = PFU × Fator de diluição × 10.
Para a realização da expansão da biblioteca CoronaScan, foi necessitado uma concentração de 1×106 de fagos para cada 10 mL das células hospedeiras com OD600 = 0,6 a 1,0 u.a. Foi separado, em diferentes tubos, 1 mL da mistura fagos/hospedeiro e adicionado 10 mL de meio top agarose aquecido a 45° C, sendo plaqueada em placas de Petri contendo
uma camada de ágar LB com ampicilina a 50 µg/mL, seguido pela incubação em TA em uma superfície plana overnight. Após a conferência da quase confluência da placa, 10 mL do tampão de extração (20 mM Tris-HCl pH 8.0, 100 mM NaCl e 6 mM MgSO4) foi adicionado e as placas foram incubadas a 4° C em uma superfície plana overnight. Assim, o tampão de extração foi transferido para um único tubo estéril e 500 µL de clorofórmio foi adicionado para uma centrifugação de 3000 g por 5 minutos, sendo a nova titulação conferida após o término. A biblioteca foi aliquotada e armazenada a -80° C em DMSO 10% (vol/vol).
A quantificação de IgG foi realizada por um ELISA sanduíche, onde uma placa de 96 poços foi adsorvida com 2 µg/mL da porção Fab de anti-IgG humana (SouthernBiotech®, Birmingham, Alabama, EUA), produzido em cabra, diluído em tampão carbonato- bicarbonato (pH 9.5) em volume final de 50 µL/poço. A placa foi selada e incubada overnight a 4ºC e, então, o anticorpo de captura não adsorvido foi descartado. Logo, a placa foi lavada com tampão de lavagem (PBS 0,05% Tween-20) e bloqueada com solução de bloqueio (PBS 5% leite desnatado Molico®, Vevey, Suíça) por 1 hora a 37° C. A placa foi lavada novamente e 50 µL/poço da solução padrão de IgG (Human IgG Normal, Invitrogen®, Waltham, Massachusetts, EUA) diluída em solução de bloqueio foi adicionada em duplicata nas concentrações de 200, 100, 33.33, 11.11, 3.7, 1.23 e 0.41 ng/mL. Concomitantemente, um soro pertencente ao banco de soros do Laboratório de Imunobiotecnologia do Instituto Adolfo Lutz (autorizada, sob o n° 32264120.5.2001.0061, no Comitê de Ética do Instituto de Infectologia Emílio Ribas) foi diluído em solução bloqueio nas razões de 1/10.000, 1/100.000, 1/400.000, 1/800.000, 1/1.000.000, 1/2.000.000, 1/4.000.000 e 1/8.000.000 e
adicionada à placa em duplicata em volume de 50 µL/poço, sendo a placa selada e incubada por 1 hora a 37° C. Em seguida, a placa foi lavada 5x para remoção do material não ligado. Posteriormente, foi feita a incubação com 50 µL/poço de uma solução de anticorpo anti- human IgG -HRP (1:5000) em TA por 30 min. Após a lavagem, 50 µL/poço do substrato TMB (3,3′,5,5′-tetrametilbenzidina) foi adicionado em TA e a placa foi coberta em papel alumínio, sendo a intensidade de coloração monitorada a cada 2 minutos. A solução de (0,2 N de H2SO4) foi adicionada seguindo a ordem de adição de TMB (50 µL/poço). Por fim, a absorbância da placa foi lida no comprimento de onda (λ) de 450 nm. Os dados obtidos foram analisados no software GraphPrism versão 8.0.1 (GraphPad Software, San Diego, Califórnia EUA). A concentração da amostra de soro foi determinada pela interpolação dos valores de absorbância obtidos na diluição 1/1.000.000 com a curva padrão pelo modelo “sigmoidal, 4 PL and X is logX (concentration)”, sendo a curva padrão da amostra de soro construída por meio da análise de regressão não-linear pelo modelo “one-site binding model”.
RESULTADOS E DISCUSSÃO
Inicialmente, a biblioteca CoronaScan, que já se encontrava em posse do Laboratório de Imunobiotecnologia do IAL, obteve uma média de titulação (realizada em duplicata) de 1,5×105 (± 0,7) pfu/mL, como demonstrado na tabela 1.

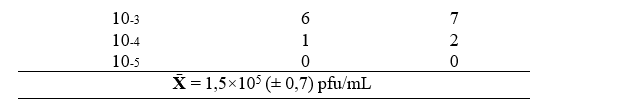
O processo morfogênico do fago T7 garante a expressão de uma maior diversidade de aminoácidos quando comparado ao fago M13, acarretando um maior número de peptídeos funcionais, a depender do quanto a nova molécula pode interferir na instabilidade da partícula viral bem como impedir a replicação do mesmo (Krumpe et al., 2007).
O método de escolha para a amplificação da biblioteca foi por ensaio de lise em placa, uma vez que o total de recombinantes primários era inferior a 5×106 pfu/mL, como demanda o protocolo da Novagen T7Select® System Manual (NOVAGEN, 2011). Após a expansão, foi possível observar pela grande confluência das placas utilizadas no procedimento que a titulação havia aumentado. Após a realização de uma nova titulação, determinou-se que a biblioteca foi aumentada cerca de 100.000 vezes, estando estimada em um título de 1,5×1010 (± 2,12) pfu/mL, conforme observado na tabela 2.

Embora o título da biblioteca tenha aumentado consideravelmente, é preocupante a qualidade da biblioteca, no que se diz a respeito à diversidade viral em que ela representa. O sequenciamento de Sanger e o NGS são duas alternativas que podem auxiliar na análise representativa dos fagos ali presentes. O sequenciamento de Sanger possibilita a investigação acerca da capacidade de síntese de oligonucleotídeos dos fagos, sendo limitado a 103 sequências, enquanto o NGS permite identificar a distribuição da frequência clonal da biblioteca e de sua integridade em uma janela de 106 a 108 ligantes, o que minimiza a subestimação da população de fagos e, consequentemente, na qualidade geral da biblioteca. Além disso, mutações e proteínas truncadas são características conhecidas por alterar a cinética de crescimento dos fagos que as apresentam, levando à proliferação vantajosa frente aos demais (Matochko & Derda, 2015; Mohan et al., 2018).
O PhIP-Seq exige, ao menos, 1 µL de soro por paciente, uma vez que apenas 2 µg de IgG para a técnica de imunoprecipitação é requisitado. Jollift e colaboradores (1982) observaram por nefelometria que os valores de referência de IgG em adultos permeiam entre 6,39 a 13,49 g/L, em uma média de 9,94 g/L. A curva padrão foi construída com os valores da concentração de IgG padrão conhecida e transformados em valores de logaritmo na base
10. A interpolação das amostras à curva padrão se mostrou ter um ajuste adequado para a curva (R2=0,99) de acordo com o software GraphPad. Assim sendo, a diluição do soro na razão de 1/1.000.000 recomendada por Mohan et al. (2018) foi adequada para a obtenção de 9 mg/mL de IgG a partir da amostra de soro, ambas as curvas podem ser observadas na figura 3.
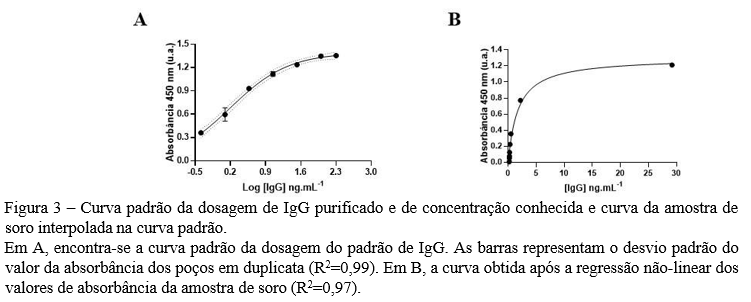
Ao utilizarem a biblioteca VirScan, uma biblioteca similar que engloba todo o viroma humano, Xu et al. (2015) constataram uma sensibilidade e especificidade de 95% quando testada e comparada para o vírus da imunodeficiência humana (HIV) e vírus da hepatite C (HCV) e seus respectivos ensaios de ELISA, assim como em teste sorológico para Herpes Simplex Virus tipo 1 (HSV-1) e tipo 2 (HSV-2), possibilitando a avaliação desses parâmetros para os vírus dos mais diversos tamanhos. Shrock e colaboradores e (2020) com a mesma técnica, descreveram uma sensibilidade de 99% e especificidade de
98% para um modelo computacional capaz de identificar a exposição contra o SARS-CoV2. Chen e colaboradores (2022) descreveram um modelo computacional, BEER, capaz de aumentar a sensibilidade de peptídeos fracamente reativos para o uso do PhIP-Seq.
Os mecanismos que desencadeiam sequelas pós-agudas do SARS-CoV-2 (PASC, do inglês Post-Acute Sequelae of SARS-CoV-2) ainda não estão muito bem esclarecidos. Uma saída para elucidar os biomarcadores que conduzem a essa condição é o uso da tecnologia PhiP-Seq, que pode ser relevante para a obtenção de respostas e questionamentos científicos, principalmente dentro do âmbito de estudos epidemiológicos de soroprevalênca (Mantovani et al., 2022).
CONCLUSÕES
Com a otimização da titulação e da expansão da biblioteca CoronaScan, a manutenção, isso é, a constante renovação da biblioteca, poderá ser feita ao longo do tempo a medida em que os ensaios de PhIP-Seq forem sendo realizados, se tornando uma fonte inesgotável de reagente. No entanto, a quantificação de IgG pelo método de ELISA ainda precisa ser aprimorada para assegurar a reprodutibilidade em amostras de soro de pacientes de diferentes quadros clínicos. Sobretudo, recomenda-se a realização do controle de qualidade de biblioteca, devido ao fato da mesma ter sido expandida a partir de um estoque cuja titulação era abaixo do valor esperado.
AGRADECIMENTOS
Aos meus colegas de laboratório de Imunobiotecnologia, que contribuíram imensamente para a construção desse trabalho e para a minha formação e a coordenadoria e professores do Curso de Especialização em Vigilância Laboratorial em Saúde Pública do Instituto Adolfo Lutz, com os quais tornaram esse projeto possível.
REFERÊNCIAS
CHEN, Athena et al. Detecting and quantifying antibody reactivity in PhIP-Seq data with BEER. Bioinformatics, v. 38, n. 19, p. 4647-4649, 2022.
DENG, Xiangying et al. Advances in the T7 phage display system. Molecular medicine reports, v. 17, n. 1, p. 714-720, 2018.
HERSTEIN, Jocelyn J. et al. Emergency preparedness: What is the future?. Antimicrobial Stewardship & Healthcare Epidemiology, v. 1, n. 1, p. e29, 2021.
JOLLIFT, C. R. et al. Reference intervals for serum IgG, IgA, IgM, C3, and C4 as determined by rate nephelometry. Clinical chemistry, v. 28, n. 1, p. 126-128, 1982. KRUMPE, L. R. H.; MORI, T. T7 lytic phage-displayed peptide libraries: Construction and diversity characterization. Methods in Molecular Biology, v. 1088, p. 51–66, 2014.
KRUMPE, Lauren RH et al. Trinucleotide cassettes increase diversity of T7 phagedisplayed peptide library. BMC biotechnology, v. 7, n. 1, p. 1-8, 2007.
MANTOVANI, Alberto et al. Long Covid: where we stand and challenges ahead. Cell Death & Differentiation, v. 29, n. 10, p. 1891-1900, 2022.
MATOCHKO, Wadim L.; DERDA, Ratmir. Next-generation sequencing of phagedisplayed peptide libraries. Peptide Libraries, p. 249-266, 2015.
MOHAN, Divya et al. PhIP-Seq characterization of serum antibodies using oligonucleotide- encoded peptidomes. Nature protocols, v. 13, n. 9, p. 1958-1978, 2018. MORGENLANDER, William R. et al. Antibody responses to endemic coronaviruses modulate COVID-19 convalescent plasma functionality. The Journal of clinical investigation, v. 131, n. 7, 2021.
NOVAGEN. T7Select® System Manual. Disponível em:
www.merck4biosciences.com.
PENG, Xue-Liang et al. Advances in the design and development of SARS-CoV-2 vaccines.
Military Medical Research, v. 8, n. 1, p. 1-31, 2021.
RAHMAN, Sayeeda et al. Epidemiology, pathogenesis, clinical presentations, diagnosis and treatment of COVID-19: a review of current evidence. Expert review of clinical pharmacology, v. 14, n. 5, p. 601-621, 2021.
SHROCK, Ellen et al. Viral epitope profiling of COVID-19 patients reveals crossreactivity and correlates of severity. Science, v. 370, n. 6520, p. eabd4250, 2020. TAN, Yuyu et al. Advance in phage display technology for bioanalysis. Biotechnology journal, v. 11, n. 6, p. 732-745, 2016.
WHO COVID-19 Dashboard. Geneva: World Health Organization, 2020. Disponível em:
https://covid19.who.int/. Acessado em: 01/12/2022.
WIEGAND, Tanner et al. The Rise and Fall of SARS-CoV-2 Variants and Ongoing Diversification of Omicron. Viruses, v. 14, n. 9, p. 2009, 2022.
XU, George J. et al. Comprehensive serological profiling of human populations using a synthetic human virome. Science, v. 348, n. 6239, p. aaa0698, 2015.
YUE, Hui et al. T7 Phage as an Emerging Nanobiomaterial with Genetically Tunable Target Specificity. Advanced Science, v. 9, n. 4, p. 2103645, 2022.
Capítulo de livro publicado no Livro da IV Mostra dos Trabalhos de Conclusão de Curso da Especialização em Vigilância Laboratorial em Saúde Pública. Para acessa-lo clique aqui.
doi.org/10.53934/9786599965821-01
Este trabalho foi escrito por:
Julia de Jesus Brito¹; Sandra Lorente²;
¹Estudante do Curso de Especialização em Vigilância Laboratorial em Saúde Pública | Programa de Citologia Oncótica – NAP – Instituto Adolfo Lutz; E-mail: [email protected]
²Docente/Pesquisador do Depto de Citologia Oncótica – NAP – Instituto Adolfo Lutz
Resumo: O Papilomavírus Humano (HPV) representa um grupo de diversos tipos de vírus
que infectam células epiteliais mucosas e cutâneas. Com base na probabilidade de desencadear o desenvolvimento de câncer, os HPVs que infectam as mucosas são classificados em alto ou baixo risco oncogênico. Os genótipos de baixo risco abrangem a
maior parte dos 200 tipos conhecidos e estão mais associados a lesões hiperproliferativas
benignas, entretanto, lesões pré-malignas, condilomas, papilomatoses e queratoses podem
estar associadas a estes tipos de genótipos. Desta forma, o objetivo deste estudo foi avaliar a associação dos vírus HPV de baixo risco oncogênico com lesões precursoras do colo uterino por meio de dados sobre a prevalência dos diferentes tipos de HPV de baixo risco em anormalidades citológicas. O estudo foi uma revisão de literatura do tipo narrativa, em que foram utilizados os dados obtidos do HPV Centre para análise da prevalência. Embora alguns genótipos de HPV de baixo risco estejam associados a lesões precursoras, este vírus parece não estar associado ao desenvolvimento do câncer de colo uterino. Através dos levantamentos realizados, concluiu-se que os HPVs de baixo-risco não são a principal causa
de lesões, no entanto, pode estar associado em lesões de alto e baixo grau, além de aparecer como principal agente causador em outras patologias.
Palavras-chave: Câncer de Colo de Útero; Papilomavírus Humano; Genótipos.
INTRODUÇÃO
O papilomavírus humano (HPV) faz parte de um grupo de diversos vírus que pertencem à família Papillomaviridae, considerada a mais bem sucedida família de vírus que infectam vertebrados, uma vez que infectam células que ainda possuem a capacidade de divisão celular, como as células basais e células metaplásicas imaturas do colo uterino, localizadas na junção escamocolunar (JEC), onde se instalam através de micro traumas no epitélio; os HPVs podem permanecer por um longo período na forma epissomal com um baixo número de cópias, não comprometendo a morfologia celular nesta fase. Existem cinco gêneros principais de HPV: alfa-papilomavirus, beta-papilomavirus, gama-papilomavirus, mu-papilomavirus e nu-papilomavírus; sendo que a infecção das células epiteliais da mucosa genital ocorre principalmente pelos 000 do tipo alfa (COHEN et al., 2019; EGAWA; DOORBAR, 2017).
O genoma dos HPVs é constituído por DNA circular de dupla fita, com tamanhos próximos a 8.000 pares de bases, caracterizado por genes que codificam as proteínas
precoces (E1, E2, E4, E5, E6 e E7) e dois genes que codificam as proteínas tardias (L1 e L2) (BZHALAVA et al., 2013; VAN DOORSLAER, 2013). Com base na probabilidade de desencadear o desenvolvimento do câncer de colo do útero, os HPVs são classificados em alto ou baixo risco oncogênico. A associação de tipos de HPV de alto risco com o câncer do colo do útero já está estabelecida, e forneceu uma justificativa para a introdução do teste DNA-HPV na triagem desta neoplasia, bem como o desenvolvimento de vacinas profiláticas contra o HPV 16 e 18, que são os principais tipos responsáveis pela doença. Outros tipos de HPV de alto risco são 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 68, 73, 82. Os HPVs de baixo risco abrangem a maior parte dos mais de 200 papilomavírus conhecidos, podem causar lesões hiperproliferativas benignas e frequentemente não causam carcinoma maligno. Os tipos 6 e 11 estão relacionados a aproximadamente 90% das verrugas genitais, e as vacinas quadrivalente e nonavalentes protegem contra a infecção por estes dois tipos. Outros tipos importantes de HPV de baixo risco são 40, 42, 43, 44, 54, 61, 70, 72, 81 (EGAWA; DOORBAR, 2017; LIU et al., 2021) Em populações susceptíveis os HPV de baixo risco podem ser refratários e causar doenças como Papilomatose Respiratória Recorrente (RRP) e Epidermodisplasia Verruciforme (EV), condições que podem se tornar fatores de risco para o desenvolvimento de câncer (EKSTRÖM et al., 2013; SCHIFFMAN et al., 2016).
As comparações funcionais dos tipos de HPV de baixo e alto risco estão longe de estarem completas, no entanto, suas principais diferenças até o momento concentram-se predominantemente nas funções exercidas pelos genes E6 e E7 codificadas por tipos de HPV de baixo risco, que têm um efeito limitado sobre os alvos celulares e as vias envolvidas no desenvolvimento de lesões e canceres. Os HPVs tipo alfa de baixo risco não induzem a desregulação completa dos genes e mecanismos da célula hospedeira. Desta maneira, o transcrito do gene E6 de HPVs de baixo risco liga-se fracamente a proteína supressora de tumor p53, que tem uma fraca inibição da resposta imunológica através de interferon, e não ativa telomerase e nem induz instabilidade genômica no hospedeiro através da proteína E7, como ocorre nos HPVs de alto risco (EGAWA; DOORBAR, 2017; SCHIFFMAN et al., 2016; THOMSEN et al., 2014).
Segundo a nomenclatura brasileira baseada no Sistema Bethesda, as anormalidades citológicas em células escamosas devem ser classificadas em atipias de células escamosas de significado indeterminado (ASC-US), lesão intraepitelial de baixo grau (LSIL), atipias de células escamosas não se pode excluir lesão de alto grau (ASC-H), lesão intraepitelial de alto grau (HSIL) e carcinoma de células escamosas. Como o Papilomavírus Humano também atinge as células glandulares que se encontram mais profundamente no canal endocervical, as anormalidades citológicas detectadas nestas células, são classificadas pelo Sistema Bethesda em células glandulares atípicas (AGC), subdividida em atipia de células glandulares endocervicais sem outras especificações (AGC-SOE) e atipias glandulares endocervicais favorecendo neoplasias (AGC-FN) (GLOBAL CANCER OBSERVATORY, [s.d.]; INSTITUTO NACIONAL DE CÂNCER JOSÉ ALENCAR GOMES DA SILVA (INCA); MINISTÉRIO DA SAÚDE, 2016; NAYAR; WILBUR, 2015).
Desta forma, para melhor compreensão do papel de genótipos de HPVs de baixo risco oncogênico no desenvolvimento de lesões precursoras do câncer do colo uterino, o objetivo deste estudo foi avaliar a associação dos vírus HPV de baixo risco oncogênico com lesões precursoras do colo uterino por meio de dados sobre a prevalência dos diferentes tipos de HPV de baixo risco em anormalidades citológicas.
MATERIAL E MÉTODOS
O estudo foi uma revisão de literatura do tipo narrativa, em que foram utilizadas as seguintes plataformas de pesquisa: PubMed, Biblioteca Virtual de Saúde (BVS), Scielo,
Lilacs e MedLine usando como descritores “Câncer de Colo de Útero”, “Condiloma”, “HPV” e suas associações na língua portuguesa e inglesa. Além disso foram realizadas pesquisas no HPV Online (https://hpvonline.com.br/), Global Cancer Observatory (https://gco.iarc.fr/), um livro e dois manuais. Na pesquisa foram obtidos 57 artigos e destes, 23 foram selecionados para a elaboração do manuscrito.
RESULTADOS E DISCUSSÃO
HPVs de baixo risco são vírus que no geral não estão envolvidos na carcinogênese do colo uterino, entretanto dados da literatura mostram lesões precursoras de câncer associadas a estes tipos de HPVs. A prevalência de HPV de baixo risco é consideravelmente menor na população feminina quando comparada ao HPV de alto risco em casos com anormalidades citológicas. No entanto, estes tipos também são detectados em lesões precursoras do câncer do colo uterino.
De acordo com os dados globais coletados através do site de domínio internacional HPV Information Centre, os resultados de prevalência mostram que os principais genótipos de HPV de baixo risco foram os tipos 6, 11, 42, 44, 62, 81 e 84 em mulheres com LSIL; 6 11,
42, 44, 54, 61, 81 e 89 em mulheres com HSIL; e os tipos 42 e 44 estavam presentes em casos de carcinoma e adenocarcinoma (HPV CENTRE, [s.d.]).
Um estudo da Dinamarca detectou a prevalência de 6,9% de HPV de baixo risco (6, 11, 42, 43 ou 44) pelo método de captura-hibrida (teste DNA-HPV) em 40.000 amostras de citologia em meio líquido. Sendo que a prevalência dos tipos HPV de baixo risco em mulheres com citologia normal foi de 6,3%, enquanto que, em mulheres com citologia classificada como ASCUS, LSIL ou HSIL teve prevalência de 33,1% 19,6% e 12,7%, respectivamente (NIELSEN et al., 2012). Martora et al (2019), na Itália, estudaram 1265 raspados pelo método de genotipagem de HPV, em mulheres entre 16 a 63 anos, que apresentaram anormalidades citológicas, e observaram uma prevalência de 12,4% de HPV 53; 9,9% de HPV 62; 9,4% de HPV 42; 8,2% de HPV 73; e 7,1% de HPV 54 (MARTORA et al., 2019).
Apesar do HPV de baixo risco estar presente em muitos casos de mulheres com anormalidades citológicas, Thomsen et al (2014) observou que o risco de neoplasia intraepitelial cervical 3, num período de 8 anos, foi de 1,7% em mulheres com HPV de baixo risco sem HPV de alto risco concomitante. Este estudo aponta que o HPV de baixo risco não prediz lesão de alto grau (THOMSEN et al., 2014).
A maior importância dos HPVs de baixo risco se dá por sua presença em verrugas anogenitais (condiloma), causadas pelo HPV dos tipos 6 e 11. Como o HPV de baixo risco possui tendência a regredir sozinho, esses condilomas, quando tratados com medicamentos tópicos, retirados cirurgicamente ou são cauterizados, costumam ressurgir até que o próprio sistema imune seja capaz de eliminar o vírus, em casos raros a possibilidade de que um condiloma evolua para uma lesão significativa deve ser considerada (COHEN et al., 2019).
Alguns relatos descritos por Pujari et al. demonstraram sua presença, em especifico o HPV 42, em lesão do tipo queratose seborreica no colo do útero, uma condição extremamente rara, pois trata-se de uma lesão cutânea benigna, é semelhante a um tumor comumente observado em pacientes idosos em áreas que ficam expostas ao sol. De 7 relatos sobre a queratose seborreica no colo uterino, 3 eram positivos para o HPV de baixo risco tipo 42 como principal agente, e em outros estudos, apareceu associado a queratoses seborreicas vulvares (18 de 25 casos) (PUJARI et al., 2021).
A Papilomatose Respiratória Recorrente (RRP), uma manifestação rara, porém grave do HPV que se encontra alojado na laringe. Uma pesquisa publicada por OUDA et al. sugere que a infecção possui três picos de idade específicos para um maior potencial de atuação, 7,
35 e 64 anos, o tumor apresenta grande morbidade em caráter de suas lesões serem confluentes e promoverem disfonia e dispneia, os tipos mais detectados são os baixo risco 6 e 11, no entanto, em alguns casos raros, os tipos alto risco 16 e 18 podem ser encontrados em crianças, associando-se a um maior poder de malignização (NETO; CAMPOS; BASTOS, 2002; OUDA et al., 2021).
A Epidermodisplasia Verruciforme, outra doença de grandes consequências associada ao HPV de baixo risco dos tipos 5 e 8, os quais se instalam nos queratinócitos, foi descrita pela primeira vez em 1922 por Lewandowsky e Lutz, trata-se de uma condição dermatológica genética onde os pacientes apresentam uma diminuição da capacidade do sistema imune em se defender e erradicar os HPVs, o que leva a uma infecção persistente e ao aumento da possibilidade de malignidade durante o desenvolvimento dessa displasia cutânea, um fato interessante é que portadores de EV não apresentam capacidade de defesa reduzida contra outros tipos de patógenos (ASZODI et al., 2020; MYERS; KWAN; FILLMAN, 2022).
Além desses aspectos, existe uma diferença em sua composição molecular, visto que, mesmo possuindo as mesmas proteínas que o HPV de alto risco, o baixo risco não possui o mesmo potencial lesivo (BZHALAVA et al., 2013; THOMSEN et al., 2014). As proteínas virais, não atingem os principais mecanismos da célula hospedeira que auxilia na sua replicação, como exemplo as proteínas E6 e E7 do HPV baixo risco, que não influenciam diretamente na divisão das células basais, mas essas proteínas podem modular a densidade celular da camada basal ou funcionar transitoriamente em estágios específicos do ciclo de vida do HPV, como durante a formação da lesão, por consequência da diferente regulação transcricional desses genes (CHEN et al., 2019). Em um momento de reentrada do vírus no ciclo celular, as proteínas E6 e E7 proporcionam um ambiente favorável à sua replicação, porém não induz a divisão celular, resultando em uma ação tardia do promotor, em que E1 e E2 aumentam, permitindo que os produtos gênicos virais e celulares amplifiquem epissomas nucleares do HPV. Como regra geral, para os dois tipos de HPV, baixo e alto risco, pode-se dizer que eles trabalham de forma similar, mas não necessariamente exercem funções idênticas durante o ciclo de vida viral, afetando sua virulência (MARTORA et al., 2019; WENTZENSEN et al., 2016).
CONCLUSÕES
O papilomavírus humano do tipo alfa de baixo risco está presente principalmente em verrugas genitais, todavia demonstram um poder carcinogênico fraco por suas particularidades genômicas, exercendo uma influência limitada sobre os mecanismos da célula hospedeira que irá ajudá-lo a se replicar. Muitos tipos de HPV baixo risco estão presentes em mulheres que possuem citologia normal e geralmente não irão se manifestar, porém algumas vezes podem se associar a outros tipos de HPV de alto risco e serem encontrados em lesões intraepiteliais.
HPVs de baixo risco são vírus que no geral não estão envolvidos na carcinogênese do colo uterino, entretanto dados da literatura mostram lesões precursoras de câncer associadas a estes tipos de HPVs. A prevalência de HPV de baixo risco é consideravelmente menor quando comparada ao HPV de alto risco em casos com anormalidades citológicas, no entanto, estes tipos também são detectados em lesões precursoras do câncer do colo uterino.
REFERÊNCIAS
ASZODI, N. et al. Epidermodysplasia verruciformis. HautarztSpringer Medizin, , 1 jul. 2020.
BZHALAVA, D. et al. A systematic review of the prevalence of mucosal and cutaneous human papillomavirus types. Virology, v. 445, n. 1–2, p. 224–231, out. 2013.
CHEN, Z. et al. Characteristics of human papillomaviruses distribution in Guizhou Province, China. Virology Journal, v. 16, n. 1, p. 1–5, 2019.
COHEN, P. A. et al. Cervical cancer. The LancetLancet Publishing Group, , 12 jan. 2019.
EGAWA, N.; DOORBAR, J. The low-risk papillomaviruses. Virus ResearchElsevier B.V., , 2 mar. 2017.
EKSTRÖM, J. et al. Diversity of human papillomaviruses in skin lesions. Virology, v. 447, n. 1–2, p. 300–311, dez. 2013.
GLOBAL CANCER OBSERVATORY. […]. Disponível em: https://gco.iarc.fr/. Acesso em: 17 nov. 2022.
HPV CENTRE. […]. Disponível em: https://hpvcentre.net/. Acesso em: 14 out. 2022.
INSTITUTO NACIONAL DE CÂNCER JOSÉ ALENCAR GOMES DA SILVA (INCA); MINISTÉRIO DA SAÚDE. MINISTÉRIO DA SAÚDE SECRETARIA DE ATENÇÃO
À SAÚDE. Rio de Janeiro: [s.n.]. Disponível em: www.saude.gov.br/sas,.
LIU, F. et al. Association of human papillomavirus genotype distribution and cervical cytology: A Cross-sectional Study. Epidemiology and Infection, 2021.
MARTORA, F. et al. Seven years prevalence and distribution of high and low risk HPV genotypes in women living in the metropolitan area of Naples. Cancer Epidemiology, v. 63, 1 dez. 2019.
MYERS, D. J.; KWAN, E.; FILLMAN, E. P. Epidermodysplasia Verruciformis Continuing Education Activity. [s.l: s.n.]. Disponível em:
https://www.ncbi.nlm.nih.gov/books/NBK534198/?report=printable.
NAYAR, R.; WILBUR, D. C. The Bethesda System for Reporting Cervical Cytology Dee nitions, Criteria, and Explanatory Notes Third Edition. [s.l: s.n.].
NETO, C. A. DE A.; CAMPOS, R. M. C.; BASTOS, M. DE L. S. Papilomatose
Respiratória Recorrente com Disseminação Pulmonar – Relato de Dois CasosRadiol Bras. [s.l: s.n.].
NIELSEN, A. et al. The importance of low-risk HPV infection for the risk of abnormal cervical cytology/histology in more than 40 000 Danish women. Sexually Transmitted Infections, v. 88, n. 8, p. 627–632, dez. 2012.
OUDA, A. M. et al. Hpv and recurrent respiratory papillomatosis: A brief review. LifeMDPI, , 1 nov. 2021.
PUJARI, R. et al. Seborrheic keratosis-like lesion of the cervix: First report of the cytological features of a low-risk HPV 42-Associated Lesion. Acta Cytologica, v. 65, n. 5, p. 448–452, 1 set. 2021.
SCHIFFMAN, M. et al. Carcinogenic human papillomavirus infection. Nature Reviews Disease Primers, v. 2, 1 dez. 2016.
THOMSEN, L. T. et al. High-risk and low-risk human papillomavirus and the absolute risk of cervical intraepithelial neoplasia or cancer. Obstetrics and Gynecology, v. 123, n. 1, p. 57–64, 2014.
VAN DOORSLAER, K. Evolution of the Papillomaviridae. Virology, v. 445, n. 1–2, p. 11– 20, out. 2013.
WENTZENSEN, N. et al. Triage of HPV positive women in cervical cancer screening. Journal of Clinical VirologyElsevier B.V., , 1 mar. 2016.
Organizadores:
Cristiane Bonaldi Cano
Elaine Lopes de Oliveira
Jackson Andson de Medeiros
Editora: Agron Science
DOI: doi.org/10.53934/9786599965821
ISBN: 978-65-999658-2-1
Data de publicação: 23/05/2023

Capítulo de livro publicado no Congresso Brasileiro de Química dos Produtos Naturais. Para acessa-lo clique aqui.
Este trabalho foi escrito por:
Antonio Deusimar Oliveira de Sousa Junior *; Lucianne Martins Lobato ;Cyntia Airagna Fortes dos Santos ; Gerson Freitas Vieira Neto ; Carlos Alberto Araújo Costa ; Rafael Carvalho da Silva
*Antonio Deusimar Oliveira de Sousa Junior – Email: [email protected]
Resumo: Atualmente, as coberturas comestíveis são utilizadas para desempenhar o papel de proteção e conservação de frutas pós-colheita minimamente processadas e frutas climatéricas, utilizando coberturas naturais e com boa aplicação unida com resultados capazes de reduzir a utilização de plástico na embalagem, possuindo antioxidantes em sua composição para minimizar os efeitos nocivos associados à deterioração da qualidade dos alimentos. Biopolímeros como proteínas, polissacarídeos, ceras e resinas são utilizados na produção desses materiais e podem ser utilizados isoladamente ou em combinação. Tendo isso em vista, o objetivo do trabalho foi reunir informações sobre o revestimento comestível para frutas climatéricas pós-colheita para contribuir com a importância de conservar frutas em regiões de temperaturas elevadas. Com base no Painel Intergovernamental sobre Mudanças Climáticas, refletindo essencialmente o padrão histórico de uma determinada variável como temperatura, vento, pressão atmosférica, umidade e precipitação. Além de diversos fatores podem influenciar as perdas pós-colheita, dentre os mais importantes destacam-se as injúrias mecânicas provocadas por embalagens inadequadas e manuseio incorreto, que geralmente começam na propriedade rural.
Palavras–chave: biopolímeros; coberturas comestíveis; revestimentos; fruticultura
Abstract: Currently, edible coatings are used to protect and preserve minimally processed post-harvest fruits and climacteric fruits, using natural coatings and with good application combined with results capable of reducing the use of plastic in packaging, having antioxidants in their composition to minimize harmful effects associated with deteriorating food quality. Biopolymers such as proteins, polysaccharides, waxes and resins are used in the production of these materials and can be used alone or in combination. With this in mind, the objective of this work was to gather information about the edible coating for post-harvest climacteric fruits to contribute to the importance of preserving fruits in regions of high temperatures. Based on the Intergovernmental Panel on Climate Change, essentially reflecting the historical pattern of a given variable such as temperature, wind, atmospheric pressure, humidity and precipitation. In addition to several factors that can influence post-harvest losses, among the most important are mechanical injuries caused by inadequate packaging and incorrect handling, which usually start on the rural property.
Key Word: biopolymers; edible toppings; coatings; fruit growing
INTRODUÇÃO
Recentemente, coberturas comestíveis são produzidas e contêm antioxidantes em sua composição para minimizar os efeitos nocivos associados à deterioração da qualidade dos alimentos e substâncias provenientes da fauna e flora brasileira e outras substâncias naturais (1).
As coberturas comestíveis podem ser utilizadas como estratégia para manter a qualidade e prolongar a vida útil de frutas e hortaliças após a colheita. Estratégias definidas como uma fina camada de materiais biológicos que se formam sob a superfície da fruta. Essas coberturas geralmente são feitas de materiais de qualidade alimentar, adicionando ou substituindo ceras que ocorrem naturalmente na superfície da fruta. Os ingredientes presentes podem ser consumidos dependendo da fruta em questão, por isso devem ser GRAS: geralmente reconhecidos como seguros para consumo humano (2).
Idealmente, esses revestimentos devem ser invisíveis, duráveis e não tóxicos e utilizados como parte dos alimentos, reduzindo as trocas gasosas e a perda de massa e atuando como uma barreira ao ataque microbiano à umidade (3).
Os filmes comumente usados são polietileno e polipropileno. Apesar da eficácia desses polímeros, razões ambientais ligadas às novas tendências têm trazido oportunidades mais sustentáveis para a indústria alimentícia (4).
Devido aos ingredientes alimentares funcionais, a demanda pública por frutas e vegetais no mercado está aumentando com dificuldades. A demanda por hortaliças e frutas minimamente processadas tem aumentado significativamente devido ao seu conteúdo nutricional, fenólicos e antioxidantes, que têm sido associados à prevenção de vários tipos de câncer e doenças degenerativas. Frutas e legumes têm uma vida útil curta devido à sua deterioração. Aproximadamente 30% das frutas e hortaliças sofrem ou se deterioram durante o transporte e manuseio devido a insetos, microrganismos, situações de pré-colheita e pós-colheita (4).
Frutas e hortaliças colhidas requerem métodos de manejo pós-colheita adequados e aprimorados para minimizar as perdas de qualidade e quantidade durante esse período (5). Durante o processo de amadurecimento, as frutas sofrem diversas alterações, como mudanças na cor, sabor, textura e aroma, que afetam diretamente aspectos importantes para a comercialização e indústria, como vida de prateleira pós-colheita, apelo visual, recepção sensorial, rendimento, suscetibilidade a ataques de microrganismos e danos mecânicos (6).
Os frutos se comportam de maneira diferente durante o amadurecimento de acordo com a síntese e respiração de etileno, o que os divide em dois grupos: climatéricos e não climatéricos. Frutos climatéricos são caracterizados por aumento súbito da respiração e síntese de etileno durante o desenvolvimento. As frutas de clima geralmente são colhidas quando os frutos ainda estão verdes. O objetivo é facilitar seu uso e prolongar sua vida útil. Frutos não climáticos não têm esse aumento na taxa de respiração e na produção de etileno (7).
O método de aplicação desse revestimento é importante para a boa conservação dos alimentos frescos, sendo necessário avaliar corretamente qual método funciona melhor variando o tipo de fruta e em quais pontos o revestimento deve funcionar. A interação entre a fruta e o revestimento é chamada de molhabilidade e é usada para avaliar sua eficácia (8).
Com o desenvolvimento da tecnologia, já é possível produzir embalagens ativas que agem como um filme ou revestimento comestível. A produção dessas embalagens tem como base: 1) um biopolímero, que se decompõe rapidamente e sem deixar resíduos, 2) plastificantes que promovem a conformabilidade e 3) aditivos que interagem com os alimentos para proteger ou retardar os efeitos de elementos como oxigênio, umidade, radiação, radiação ultravioleta, microrganismos, etileno e outras substâncias nocivas que fazem parte da degradação dos alimentos e afetam a vida útil (9, 10).
Nesse contexto, os revestimentos comestíveis parecem ser uma opção, pois são derivados de proteínas, polissacarídeos, lipídeos e compostos. Entre as proteínas utilizadas para fazer coberturas comestíveis, destacam-se o amido e a caseína. Devido à sua abundância e baixo custo, o amido tem sido amplamente utilizado para desenvolver revestimentos comestíveis. Além disso, os revestimentos de amido de alimentos são transparentes, incolores, inodoros e têm baixa permeabilidade ao oxigênio (11).
Os estudos de Dantas et al. (2015), experimentou a incorporação dos revestimentos na preservação das frutas laranja e maracujá, alterando propriedades mecânicas, térmicas e permeáveis, devido a quantidade de fibras presentes nas frutas.
A utilização de revestimentos alimentícios para preservação pós-colheita de frutas inteiras ou minimamente processadas tem sido considerada uma tecnologia emergente com alto potencial econômico, principalmente para frutas tropicais e amplamente exportadas (12). Dessa forma, o objetivo do trabalho foi reunir informações sobre o revestimento comestível para frutas climatéricas pós-colheita, em detrimento dos fatores que alteram a qualidade do fruto em relação ao clima da região predominante.
TÓPICOS
Cenário de frutíferas climatéricas no Brasil
A produção de frutas no Brasil se estabiliza na margem de produção em 41 milhões de toneladas, com ocupação de 2,6 milhões de hectares, ou seja, 0,3% do território nacional pertence à produção de fruticultura, em frente às lavouras que ocupam 7,8%. O país possui em torno de 940 mil propriedades agropecuárias distribuídas em todas as regiões do país, onde 81% se enquadram como agricultura familiar. Devido à grande demanda no setor frutícola, o setor disponibilizou emprego registrado para 193,9 mil pessoas, aumento de 9% em relação ao ano de 2020 (13).
Através dos portos de Pecém e Mucuripe, o estado do Ceará se fortaleceu nacionalmente como uma força na agricultura irrigada de alta tecnologia, na produção de frutas, hortaliças e flores para o mercado interno e para exportação. São as vantagens competitivas de localização que têm estimulado o emprego e a renda no meio rural e melhorado a balança comercial. Considerando os dez primeiros meses de 2020, o Ceará exportou 17,2% mais frutas em relação ao mesmo período do ano passado (US $50,2 milhões contra US $2,8 milhões). Os destaques das exportações são frutas como melão, banana, melancia, manga, mamão, caju, coco, maracujá e acerola. (14).
Em 2021, a exportação de frutas frescas do Brasil atingiu recorde em volume e faturamento, apoiada pelo desenvolvimento comercial e produtivo do setor de fruticultura, apesar de bastante otimista, o resultado final das entregas em 2022 não vai manter o recorde alcançado no ano passado. As principais razões para o resultado mais fraco em 2022 são problemas logísticos (falta de contêineres e aumento do valor do frete, tanto marítimo quanto aéreo), altos custos de produção (reforçados pela guerra entre Rússia e Ucrânia, que elevou os valores) , margens reduzidas para exportadores em insumos, principalmente fertilizantes e produção reduzida de algumas frutas em 2022 devido ao clima desfavorável (15).
De acordo com o Instituto Brasileiro de Geografia e Estatística (IBGE) as frutas têm uma produção com crescimento acentuado no mercado interno e nas exportações no Brasil, principalmente no Período de Janeiro a abril de 2021, Pesquisas apontam em torno de 450 mil toneladas de frutas gerando US$ 422,3 milhões de receita ao país, principalmente no estado de São Paulo. Dessa forma, os fruticultores e especialistas asseguram que a exportação é uma atividade que estimula o mercado interno e beneficia a rentabilidade com a produção (16).
Ainda com base nessa projeção de frutas do país, o setor de fruticultura do Brasil bateu recorde, chegando a US $1 bilhão em exportações de frutas em 2021 (16). O país enviou cerca de 1,2 milhão de toneladas de frutas para o mercado internacional, 18% a mais que no ano passado, o faturamento foi de 1,06 bilhão de dólares, crescendo 20% na mesma comparação. De acordo com a Associação Brasileira dos Produtores e Exportadores de Frutas e Derivados (16).
A mudança climática é um fator limitante na armazenagem pós colheita de frutas no Brasil, pois consiste numa alteração a longo prazo no clima global ou regional. Com base no Painel Intergovernamental sobre Mudanças Climáticas, principalmente em reflexo ao padrão histórico de determinada variável como temperatura, ventos, pressão atmosférica, umidade do ar e chuva (17).
Associado com a mudança climática, outros fatores são decisivos na qualidade de um produto natural, principalmente o armazenamento e a comercialização. Em vista disso, a aplicação de coberturas e revestimentos comestíveis protetores tem se apresentado como técnica útil na preservação de produtos minimamente processados com vida útil reduzida, devido aos processos pelos quais são submetidos (18).
A embalagem consiste na tecnologia utilizada para entregar produtos seguros aos consumidores finais a um preço acessível. Na preservação de alimentos, tal tecnologia é extremamente necessária, pois permite a comercialização de produtos com maior resistência ao tempo diferente do local de produção, e aumenta a durabilidade em transporte (19).
Revestimento comestível
O crescente volume de utilização de materiais plásticos e as implicações ambientais inerentes ao seu descarte não racional pós-consumo, como no setor de alimentos, tem preocupado a sociedade. Tradicionalmente, os plásticos são derivados do petróleo, caracterizando-se por ser um material não biodegradável e inerte, uma vez que é sabido que pode levar algumas dezenas ou centenas de anos para se degradar no meio ambiente (12). Contudo, são materiais com grande aplicabilidade e versatilidade, em geral são resistentes, leves, e o que mais chama a atenção é que são de baixo custo, o que os torna bastante atrativos para um grande número de aplicações que podem variar desde fabricação de peças à produção de embalagens (12).
Definidos de acordo com suas características de aplicação, filmes comestíveis são estruturas pré-formadas originadas por moldagem de compressão, fundição ou extrusão, geralmente utilizados para acondicionar produtos finais (20); já os revestimentos são formados por uma película pela imersão, pulverização ou solução dispersa na superfície do alimento (21,22). Suas propriedades de formação de filme permitem a síntese de membranas (espessura > 30μm) e revestimentos (< 30 μm) que são utilizados com sucesso para a conservação dos alimentos (23).
Filmes produzidos com polímeros biodegradáveis oferecem uma alternativa sustentável utilizando embalagens menos prejudiciais ao meio ambiente, além de aumentar a vida da prateleira de alimentos. O desenvolvimento de materiais biodegradáveis a partir de fontes renováveis e a consequente redução no uso de polímeros plásticos sintéticos reduziram os impactos ambientais esperados pela indústria de embalagens (24).
Para a produção desses materiais, são usados biopolímeros como proteínas, polissacarídeos, ceras e resinas no preparo de revestimentos, podendo ser empregados puros ou em combinação (25). O uso dos revestimentos comestíveis traz benefícios aos frutos da prateleira tanto no prolongamento da vida útil, quanto atratividade ao consumidor, conservando o alimento, pois reduz sua taxa de respiração e retarda a perda de compostos voláteis.
Villadiego e colaboradores (9) definem os revestimentos comestíveis como biopolímeros de origem animal ou vegetal, aplicados diretamente sobre a superfície do alimento, que após secos formam uma fina camada que age como uma barreira protetora aos elementos externos, como umidade, óleo e vapor orgânico, estendendo a vida de prateleira, sem comprometer os aspectos sensoriais como cor, nem sabor.
Entre os biopolímeros utilizados em revestimentos comestíveis, os amidos são os que têm mostrado eficiência. O amido, depois da celulose, é o mais abundante no planeta e consiste de moléculas lineares (amilose) e ramificadas (amilopectina) de glicose, e é amplamente usado pela sua versatilidade, abundância, baixo custo e propriedades protetivas benéficas na conservação de frutas e hortaliças (26).
A quitosana é um polissacarídeo natural, barato e disponível comercialmente. Além da sua versatilidade, esse material detém outras propriedades interessantes comercialmente, tais como: atoxicidade, renovabilidade, biodegradabilidade, biocompatibilidade, além de propriedades antibacterianas, antifúngicas e de afinidade com proteínas (27).
A quitosana é obtida a partir do exoesqueleto de crustáceos (camarão, caranguejo de garra vermelha e conchas de caranguejo), paredes celulares de fungos e outros materiais biológicos, sendo um dos poucos polissacarídeos catiônicos e que exibe as propriedades químicas desejadas para aplicações em larga escala em vários campos. Por ser é atóxica e biodegradável no corpo humano, a quitosana é um material muito atrativo para RCs (revestimento comestíveis) devido às suas propriedades antimicrobianas contra bactérias, fungos e leveduras (28, 29).
Revestimentos comestíveis podem ser preparados e aplicados de três formas diferentes: pulverização, imersão ou por espalhamento (30). A pulverização se dá utilizando métodos de pulverização tradicionais ou por “eletro pulverização”em que é usado um aparelho que emprega eletricidade para dispersar um líquido; a imersão é geralmente utilizada em alimentos em que é necessário a aplicação de uma camada mais espessa. Esse método é utilizado de modo a melhorar a qualidade de propriedades físico-químicas. Na técnica de espalhamento, a aplicação do revestimento é feita pincelando o alimento com o revestimento.
As vantagens dos revestimentos comestíveis incluem compostos que são biodegradáveis e podem ser usados com alimentos, preservam as propriedades da fruta quando revestidas, melhoram o valor nutricional e as propriedades organolépticas da fruta por ser um meio de transporte de compostos ativos. Além disso, por serem derivados de produtos naturais encontrados na natureza, são fáceis de encontrar e não trazem custos para o fabricante (30).
Lopes et al., (31), concluíram em seu estudo em conservação de goiabas, que o uso dos revestimentos comestíveis de caseína e amido é eficiente na redução de perda de massa das goiabas. A adição do extrato de barbatimão nos revestimentos comestíveis de amido e caseína proporcionou uma maior conservação da firmeza e sua coloração, evitando seu amarelecimento e clareamento. Sendo assim temos a opção de utilização do barbatimão associado aos revestimentos comestíveis, pois apresentou potencial para uma melhor conservação de goiabas.
Segundo Costa et al., (32), pode ainda ser utilizado o revestimento comestível de amido de mandioca, que tem apresentado na última década resultados promissores na conservação de frutos e hortaliças, aumento a vida de prateleira dos produtos, garantindo os aspectos nutricionais e bioativos. Ressaltam ainda a importância de conhecer a estrutura da matéria prima para a composição da embalagem biodegradável, que deve ser compatível com os produtos que contêm, sendo sensorialmente neutros para não influenciar na qualidade sensorial dos produtos.
Os revestimentos comestíveis devem atender a vários requisitos como propriedades de barreira satisfatórias para água, gases e outras substâncias de interesse; solubilidade em água e gordura; cor e aparência adequadas; propriedades mecânicas e reológicas. Outra característica fundamental para um revestimento comestível é que não seja tóxico quando aplicado em frutas e legumes frescos, inteiros ou minimamente processados. Produtos frescos são bem sensíveis à perda de água, que causam enrugamento, perda de turgescência e deterioração da textura, por isso é importante ter condições satisfatórias para água, gases e outras substâncias de interesse (33, 34).
Mais importante ainda, os revestimentos podem ser usados com segurança em produtos embalados e são capazes de proteger os alimentos contra danos microbianos e mecânicos (35, 36, 37).
Perdas Pós-Colheita de frutíferas no Brasil
Desde a última década, o Brasil deixou de importar alimentos para se tornar um dos maiores exportadores de produtos agrícolas do mundo. Atualmente, é considerado o terceiro produtor mundial de frutas, com uma produção de mais de 40 milhões de toneladas ao ano, atrás apenas da China e Índia (38).
Em consequência disso, o país se tornou autossuficiente no abastecimento da maior parcela dos hortifrútis, garantindo fornecimento contínuo à população. No entanto, as perdas pós-colheita são expressivas, sendo estimadas em 40-50% da produção (39). Enquanto em países como os Estados Unidos as perdas não ultrapassam 10% (40). Assim, o grande volume de perdas na produção brasileira representa um desperdício de recursos, como terra, água, energia e insumos (41), e alto prejuízo econômico para o setor varejista, correspondendo em média a 600 milhões de reais por ano (42).
Em regiões tropicais como é o caso do Brasil, as perdas pós-colheita são mais acentuadas, devido às condições ambientais com temperaturas elevadas e umidade que aceleram o metabolismo dos frutos, e que aliados a ausência de uma cadeia de frio eficiente durante a comercialização, comprometem a conservação adequada do produto (42).
Diversos fatores podem influenciar as perdas pós-colheita, dentre os mais importantes destacam-se a injúrias mecânicas provocadas por embalagens inadequadas e manuseio incorreto, que geralmente começam na propriedade rural, durante a classificação e seleção dos produtos, estendendo-se até os consumidores intermediários e finais, constituindo os principais problemas da cadeia produtiva (40). Esses fatores prejudicam a comercialização das frutas, uma vez que os aspectos físicos dos produtos podem determinar os preços de compra e venda (42).
Na maioria das vezes os ferimentos gerados durante a colheita ou manipulação dos frutos tornam-se uma porta de entrada para microrganismos, como fungos e bactérias. Ao penetrarem, esses microrganismos ocasionam a podridão dos frutos, pois sua ação está relacionada à alterações nas características físicas, químicas, sensoriais e visuais, levando à perdas na produtividade e consequentemente reduzindo a vida de prateleira dos produtos (43).
Desta forma, a utilização de coberturas ou revestimento comestível surge como uma alternativa promissora para elevar a vida em pós-colheita de frutas e legumes. Essas coberturas ou filmes como também são conhecidas, não possuem a função de substituir os métodos tradicionais de conservação, como a utilização do frio ou embalagens protetoras, mas sim funcionar como um aditivo, contribuindo para melhorar a conservação dos frutos, preservando sua textura e valor nutricional, além de reduzir as trocas gasosas com o meio e as perdas ou ganho de água (44).
Taxas de exportação e importância econômica de revestimentos comestíveis
A pandemia da COVID-19 criou oportunidades para muitos empreendedores privados entrarem no mercado para atender ao aumento da demanda por alimentos. Isso, por sua vez, beneficiou os fabricantes de ingredientes alimentícios, incluindo revestimentos. A necessidade de lanches inovadores tem sido consistentemente alta nos países desenvolvidos devido ao armazenamento, utilização e entrega rápida de nutrientes e energia após o consumo. Consequentemente, tais fatores impulsionam o mercado em tempos turbulentos. As vendas de frutas e vegetais aumentaram no ano encerrado em 2020, quando surgiu a demanda por alimentos nutritivos. Portanto, apoiou o uso de filmes e coberturas comestíveis (45).
O revestimento comestível é uma tecnologia ecológica usada em muitos produtos para controlar a transferência de umidade, troca gasosa ou processos de oxidação. Uma vantagem importante do uso de filmes e revestimentos comestíveis é que vários ingredientes ativos podem ser adicionados à matriz polimérica e ingeridos com alimentos, o que aumenta a segurança ou mesmo as propriedades nutricionais e sensoriais (45).
O setor de frutas do Brasil bateu recorde ao atingir a marca de US $1 bilhão em exportações de frutas em 2021. O cenário de exportação ficou dessa forma, a manga com 272,5 mil toneladas de fruta, aumentando 12% em relação à 2020, a maçã teve crescimento de 79%, com 58% em relação a 2020. Ao todo, foram embarcadas 99 mil toneladas de manga ao mercado internacional, aproximadamente 70% dos envios das frutas brasileiras (46).
Frutas e hortaliças são alimentos frescos com grande variedade, principalmente em países tropicais como o Brasil. São alimentos ricos em nutrientes, fonte de vitaminas, carboidratos, fibras e minerais, compostos bioativos e baixo teor calórico. Estas propriedades beneficiam a saúde e desempenham um papel importante no crescimento, manutenção e desenvolvimento do corpo humano (47).
O volume total de frutas exportado em janeiro de 2021 foi de 84,91 mil toneladas, abaixo 5,06% em relação a janeiro do ano anterior, e o valor auferido foi de US $67,84 milhões, 6,5% abaixo para o mesmo período. Destaque para o crescimento do volume das exportações de melancias e mangas e queda para limões e limas, bananas, melões e mamões. Foram exportadas 8,91 mil toneladas de frutas em janeiro de 2021, 5,06% a menos que em janeiro de 2020, e o valor agregado foi de 67,8 milhões de dólares, 6,5% a menos que no mesmo período. Destaca-se o aumento das exportações de melancia e manga em e a queda das exportações de limões e limas, bananas, melões e mamões (48).
Os revestimentos preservam efetivamente a qualidade dos produtos e reduzem significativamente o uso de plástico. Além disso, esse desenvolvimento pode ser destinado ao uso de outros alimentos com efeitos conservantes ou antimicrobianos para prolongar a vida útil da fruta e juntos transportar esses efeitos benéficos ao consumidor (49).
Com isso, a indústria alimentícia pode contar com uma alternativa natural vantajosa para embalar diversos tipos de alimentos, minimizando as perdas pós-colheitas de forma segura. Esta técnica também mantém o sabor, o frescor, a aparência e a qualidade nutricional de frutas e vegetais, além de contribuir com o planeta, reduzindo o impacto ambiental que o descarte das embalagens plásticas causa ao nosso meio ambiente. (49).
CONCLUSÕES
Diversos fatores podem determinar a qualidade dos produtos naturais, dentre os mais importantes estão as propriedades organolépticas e nutricionais, bem como as condições de higiene, que na maioria das vezes estão relacionadas com o processo de armazenamento e comercialização. O uso de coberturas comestíveis tem propiciado resultados positivos, como uma prática auxiliar na conservação de produtos perecíveis, principalmente dos alimentos minimamente processados, cujo tempo de prateleira é mínimo em função dos processos pelos quais esses produtos foram submetidos.
A adesão às coberturas comestíveis ainda se encontra em fase de desenvolvimento, mas espera-se que a utilização desta tecnologia aumente significativamente nos últimos anos, em virtude do surgimento de novos biopolímeros, aumento no número de pesquisas sobre o assunto e interesse do setor produtivo e de transporte. Portanto, é esperado que o uso de revestimentos comestíveis se torne uma prática de rotina na conservação de alimentos, principalmente os de origem tropical.
REFERÊNCIAS
(1) Matta E, Tavera-quiroz MJ, Bertola N. Active edible films of methylcellulose with extracts of green apple (Granny Smith) skin. 2019; 124: 1292-1298.
(2) Burdock GA, Carabin IG. Generally recognized as safe (GRAS): history and description. Toxi Letters. 2004; 150: 3-18.
(3) Leite AR. Controle da deterioração fúngica em morangos utilizando
Extratos brutos vegetais, frente ao fungo botrytis cinerea (monografia). Campo Mourão: Universidade Tecnológica Federal do Pará; 2021.
(4) Hassan B, Chatha SAS, Hussain AI, Zia KM, Akhtar N. Recent advances on polysaccharides, lipids and protein based edible
films and coatings: A review. International Jour of Biol Macromolecules. 2018; 109: 1095-1107.
(5) Singh V, Hedayetullah M, Zaman P, Meher J. Postharvest technology of fruits and vegetables: an overview. Journal of Postharvest Technology. 2014; 02: 124-135.
(6) Magalhães DS. Desenvolvimento e maturação de frutos de pitaia vermelha de polpa branca (Dissertação). Lavras: Universidade Federal de Lavras; 2017.
(7) Barry CS, Giovannoni JJ. Ethylene and fruit ripening. Journal of Plant Growth Regulation. 2007; 26: 143-159.
(8) Parreidt TS, Schmid M, Muller K. Effect of immersion and vacuum impregnation techniques with alginate-based coating on the physical quality parameters of Cantalupo melon. Journal of Food Science. 2018; 83: 929-936.
(9) Villadiego AMD, Soares, NFF, Andrade NJ, Puschmann R, Minim VPR, Cruz R. Filmes e revestimentos comestíveis na conservação de produtos alimentícios. Revista Ceres. 2005; 52: 221–244.
(10) Paidari S, Zamindar N, Tahergorabi R, Kargar M, Ezzati S, Shirani N, Musavi SH. Edible coating and flms as promising packaging: a mini review. Journal of Food Mea and Charac. 2021; 15: 4205-4214.
(11) Yan Q, Hou H, Guo P, Dong H. Effects of extrusion and glycerol content on properties of oxidized and acetylated
corn starch-based films. Carbohydrate Polymers. 2012; 87: 707-712.
(12) FRATARI, Silvio Cesar et al. Revestimentos comestíveis para conservação pós colheita de banana: uma revisão. Verruck, S. Avanços em Ciência e Tecnologia de Alimentos, v. 4, p. 444-467, 2021.
(13) Fonseca LABV. Fruticultura Brasileira: Diversidade e sustentabilidade para alimentar o Brasil e o Mundo [Internet]. 2022 [15 de dez 2022]. Disponível em: https://cnabrasil.org.br/noticias/fruticultura-brasileira-diversidade-e-sustentabilidade-para-alimentar-o-brasil-e-o-mundo.
(14) Lima CC. Ceará se destaca como produtor e exportador de frutas [Internet]. 2020 [acesso em 01 jan 2023]. Disponível em: https://www.gazetadopovo.com.br/agronegocio/ceara-se-destaca-como-produtor-e-exportador-de-frutas/.
(15) Geraldini F, Deleo JPB, Barbieri MG, Moreira MM, Boteon M, Barros, GSC. Anuário HF Brasil: retrospectiva 2022 & perspectiva 2023 [Internet]. São Paulo: Hortifruti Brasil. 2022 [acesso em 18 dez 2022]. Disponível em: https://www.hfbrasil.org.br/br/revista/acessar/completo/anuario-hf-brasil-retrospectiva-2022-perspectiva-2023.aspx.
(16) ABAFRUTAS. IBGE projeta expansão da produção de frutas no País nos próximos anos [Internet]. 2023 [acesso em 02 jan 2023]. Disponível em: https://abrafrutas.org/2022/01/ibge-projeta-expansao-da-producao-de-frutas-no-pais-nos-proximos-anos/.
(17) Intergovernmental Panel on Climate Change – IPCC. Climate Change 2014: Synthesis Report.Genebra; 2014.
(18) Garcia DM, Dos Reis RC, Costa LM, Ferreira NMP. Uso de revestimento comestível a base de resíduo de frutas adicionado de polpa de acerola para a conservação defrutas minimamente processadas. Brazilian Jour of Dev. 2022; 8: 6301-3612.
(19) Priyadarshi R, Rhim JW. Chitosan-based biodegradable functional films for food packaging applications. Innovative Food Scie Eme Tec. 2020; 62: 102346.
(20) Khuntia A, Prasanna NS, Mitra J. Technologies for Biopolymer‐Based Films and Coatings. Biopolymer‐Based Food Packaging: Inno and Tec Appl. 2022; 3: 66-109.
(21) Cazón P, Vázquez M, Velazquez G. Cellulose-glycerol-polyvinyl alcohol composite films for food packaging: Evaluation of water adsorption, mechanical properties, light-barrier properties and transparency. Carbohydrate Polymers.
2018; 195: 432-443.
(22) Ribeiro AM, Estevinho BN, Rocha F. Preparation and Incorporation of Functional Ingredients in Edible Films and Coatings. Food Bioprocess Technol. 2021; 14: 209-231.
(23) Nair MS, Tomar M, Punia S, Kukula-Koch W, Kumar M. Enhancing the functionality of chitosan- and alginate-based active edible coatings/films for the preservation of fruits and vegetables: A review. International J of Bio Macro. 2020; 164: 304-320.
(24) Costa F, Braga RC, Bastos MSR, Dos Santos DN, Frota MM. Revestimentos comestíveis à base de fécula de mandioca (manihot esculenta) em produtos vegetais: uma revisão. Research Society and Development. 2022; 11: 1-13.
(25) Jafarzadeh S, Nafchi AM, Salehabadi A, Oladzad-abbasabadi N, Jafari SM. Application of bio-nanocomposite films and edible coatings for extending
the shelf life of fresh fruits and vegetables. Advances in C and I S. 2021; 291: 102405.
(9) Villadiego AMD, Soares, NFF, Andrade NJ, Puschmann R, Minim VPR, Cruz R. Filmes e revestimentos comestíveis na conservação de produtos alimentícios. Revista Ceres. 2005; 52: 221–244.
(26) Rocha GO, Farias MG, Carvalho CWP, Ascheri JLR, Galdeano MC. Filmes compostos biodegradáveis a base de amido de mandioca e proteína de soja. Polímeros. 2014; 24: 587-595.
(27) Bakshi OS, Selvakumar D, Kadirvelu K, Kumar NS. Chitosan as anenvironment friendly biomaterial – a review on recent modifications and applications. Int J of B Macro. 2020; 150: 1072-1083
(28) Costa MJ, Maciel LC, Teixeira JA, Vicente AA, Cerqueira MA. Use of edible films and coatings in cheese preservation: Opportunities and challenges. Food R Inter. 2018; 107: 84-92.
(29) Ortiz-Duarte G, Pérez-Cabrera LE, Artéz-Hernández F, Martínez-Hernández GB. Ag-chitosan nanocomposites in edible coatings affect the quality of fresh-cut melon. Postharvest B and Tec. 2019; 174: 174-184.
(30) Dhumal CV, Sarkar P. Composite edible films and coatings from food-grade biopolymers. J Food Sci Technol. 2018; 55: 4369-4383.
(31) Lopes AR, Dragunski DC, Caetano J, Francisco CB, Bonfim-Junior LF. Conservação de goiabas com revestimentos comestíveis de amido e caseína com extrato de barbatimão. Eng na Agricultura. 2018; 26: 295-305.
(32) Costa F, Braga RC, Bastos MSR, Dos Santos DN, Frota MM. Revestimentos comestíveis à base de fécula de mandioca (manihot esculenta) em produtos vegetais: uma revisão. Research, S and Dev. 2022; 11: 1-13.
(33) Arquelau PBF, Silva VDM, Garcia MAVT, De Araújo RLB, Fante CA. Characterization of edible coatings based on ripe “Prata” banana peel flour. Food Hydrocolloids. 2019; 89: 570-578.
(34) Chakravartula SSN, Cevoli C, Balestra F, Fabbri A, Rosa MD. Evaluation of drying of edible coating on bread using NIR spectroscopy. J of F Engi. 2019; 240: 29-37.
(35) Arnon-Rips H, Porat R, Poverenov E. Enhancement of agricultural produce quality and storability using citralbased edible coatings; the valuable effect of nano-emulsification in a solidstate delivery on fresh-cut melons model. F Chemistry. 2019; 277: 205-212.
(36) Dehghani S, Hosseini SV, Regenstein JM. Edible films and coatings in seafood preservation: A review. Food Chem. 2018; 240: 505-513.
(37) Spasojević L, Katona J, Bučko S, Savić SM, Petrović L, Milinković Budinčić J, et al. Edible water barrier films prepared from aqueous dispersions of zein nanoparticles. LWT. 2019; 109: 350-358.
(38) Kist BB, de Carvalho C, Beling RR. Anuário Brasileiro de Horti&Fruti. Santa Cruz do Sul: Gazeta; 2021.
(39) Magnoni-Junior L, Massambani O, Stevens D, da Silva WTL, do Vale JMF, Purini SRM. JC na Escola Ciência, Tecnologia e Sociedade: Mobilizar o Conhecimento para Alimentar o Brasil. São Paulo: Centro Paula Souza; 2017.
(40) Guerra AMNM, Costa ACM, Ferreia JBA, Tavares PRF, Vieira TS. Perdas pós-colheita em hortaliças provocadas por danos na rede varejista de Santarém-PA. RBAS. 2018; 8: 106-114.
(41) Spagnol WA, Silveira-junior V, Pereira E, Filho-Guimarães N. Redução de perdas nas cadeias de frutas e hortaliças pela análise da vida útil dinâmica. BJFT. 2018; 21: 1-10.
(42) Teixeira IS, Rufino MSM, Pinto CM, de Almeida AOG. Causas de perdas pós-colheita em cultivares de tomates comercializados na Ceasa, Ceará. Revista Verde. 2022; 17: 135-142.
(43) Oliveira JCF. Efeito no revestimento de própolis verde na qualidade de frutas de mamão formosa (Carica papaya L) [monografia]. Mato Grosso: Universidade Federal de Mato Grosso; 2018.
(44) Gomes IR, Resende ÉD, do Amaral DP. Uso de revestimento ativo em frutos: uma tecnologia emergente. CIAGRO. 2020; 1: 1-14.
(45) MordorIntelligence. Mercado global de filmes comestíveis e revestimentos – crescimento, tendências, impacto do covid-19 e previsões (2021 – 2026) [internet]. 2023 [acesso em 10 jan 2023]. Disponível em: https://www.mordorintelligence.com/pt/industry-reports/edible-films-and-coating-market.
(46) ABRAFRUTAS. Manga foi a fruta mais exportada pelo Brasil em 2021 [internet]. 2023 [acesso em 10 jan 2023]. Disponível em: https://abrafrutas.org/2022/01/manga-foi-a-fruta-mais-exportada-pelo-brasil-em-2021/.
(47) Silva SLR. Interferentes no consumo de frutas e hortaliças de usuários do programa
academia da saúde [Dissertação]. Belo Horizonte: Universidade Federal de Minas Gerais, Escola de Enfermagem; 2019.
(48) Companhia Nacional de Abastecimento-CONAB. Boletim Hortigranjeiro. Brasília: PROHORT; 2021. (49) Sousa A, Carvalho S, Plácido GR. Embalagens Comestíveis para frutas e vegetais: aspecto de segurança de alimentos [Internet]. 2022 [24 de fev de 2023]. Disponivel em: https://foodsafetybrazil.org/embalagens-comestiveis-para-frutas-e-vegetais-aspectos-de-seguranca-de-alimentos/.
Capítulo de livro publicado no Congresso Brasileiro de Química dos Produtos Naturais. Para acessa-lo clique aqui.
Este trabalho foi escrito por:
Isabelle Franca da Costa Vasconcelos1* ; Izabela Queiroz Silva1 ; Pedro Garcia Pereira Silva1 ; Janaína Fernandes de Medeiros Burkert1 ; Lucielen Oliveira Santos1
1 Escola de Química e Alimentos, Universidade Federal do Rio Grande – FURG
*Autor correspondente (Corresponding author) – E-mail: [email protected]
Resumo: Os carotenoides são pigmentos lipossolúveis que podem ser produzidos por via biotecnológica, como é o caso da levedura carotenogênica Rhodotorula mucilaginosa CCT 7688. Esses compostos têm importância comercial devido a sua atividade antioxidante e utilização na indústria de alimentos, destacando-se os de fontes naturais. Neste contexto, os carotenoides produzidos pela R. mucilaginosa são sintetizados no interior das células, sendo necessário realizar o processo de extração da biomassa. Desta forma, o objetivo deste trabalho foi avaliar diferentes pré-tratamentos na biomassa seguido de ruptura celular com dimetilsulfóxido (DMSO), visando maior obtenção de carotenoides e redução no tempo de processo. Para tal, diferentes tratamentos foram realizados modificando o tempo de secagem e congelamento da biomassa (24 ou 48 h) e o ciclo de ruptura com DMSO. O pré-tratamento avaliado que obteve a maior concentração específica (272,81 µg g-1) e volumétrica (1762,38 µg L-1) de carotenoides foi utilizando secagem e congelamento por 24 h, seguidos de 3 ciclos de ruptura com DMSO, sendo 36,4 % mais eficiente do que a metodologia comumente utilizada na literatura, reduzindo o tempo do processo em 48 h.
Palavras–chave: Cultivo microbiano; biomassa; extrato carotenogênico; pré-tratamento
Abstract: Carotenoids are liposoluble pigments that may be produced by biotechnology, as in the carotenogenic yeast Rhodotorula mucilaginosa CCT 7688. These compounds have commercial importance due to their antioxidant activity and use in the food industry, highlighting those from natural sources. In this context, the carotenoids produced by R. mucilaginosa are synthesized inside the cells, being necessary to carried out the extraction process. Thus, this study aimed to evaluate different pre-treatments in biomass followed by cell disruption with dimethylsulfoxide (DMSO), to obtain more carotenoids and reduce the processing time. For this purpose, different treatments were performed by modifying the drying and freezing time of the biomass (24 or 48 h) and the DMSO disruption cycle. The pretreatment that obtained the highest specific concentration (272.81 µg g-1) and volumetric concentration (1762.38 µg L-1) of carotenoids was using drying and freezing for 24 h, followed by 3 cycles of DMSO disruption, being 36,4 % more efficient than the methodology commonly used in the literature, reducing the processing time by 48 h.
Key Word: Microbial culture; biomass; carotogenic extract; pretreatment
INTRODUÇÃO
Os carotenoides são pigmentos lipossolúveis que possuem ação antioxidante natural, as células utilizam esses compostos como mecanismo de defesa sob estresse oxidativo (1–3). Eles são sintetizados por fungos, algas, plantas e bactérias, e podem ser utilizados em alimentos, como aditivo alimentar e também em medicamentos (2,4). Esses pigmentos apresentam coloração que vai do amarelo ao vermelho, sendo muitas vezes instáveis e sensíveis a luz e ao oxigênio (3,5,6). Portanto, por possuírem coloração, são utilizados como corantes naturais na indústria de alimentos e seu consumo – proveniente de produção natural – demonstra ser mais benéfico a saúde, pois são compostos não sintetizados pelo organismo humano (5,7).
A Rhodotorula mucilaginosa CCT 7688, pertencente ao grupo de leveduras oleaginosas, tem potencial de produzir carotenoides a partir de meios agroindustriais (8). Esses compostos são de importância comercial e há crescimento de sua demanda no mercado farmacêutico devido as suas benéficas atividades antioxidantes (1,2). A produção de carotenoides por cultivos microbianos apresenta vantagens, além de ser de origem natural, apresentam rápida taxa de crescimento quando comparada a outras formas de produção natural, possibilidade de cultivar durante todo o ano e produção industrial em áreas reduzidas (6,7,9).
Sabe-se que os custos de produção a partir de cultivos microbianos são altos (10). A fim de diminuir os custos do meio de cultivo, diferentes substratos, tais como alguns coprodutos agroindustriais, podem ser utilizados, como a água de maceração de milho e o melaço de cana (10,11). Esses coprodutos agroindustriais são ricos em nutrientes (carbono, nitrogênio, glicose, entre outros) (10,12,13) e podem ser utilizados como substrato por leveduras produtoras de carotenoides.
Os carotenoides produzidos pela R. mucilaginosa são sintetizados no interior das células, assim, é necessário realizar o processo de ruptura celular para fazer a recuperação (5,7,14,15), além disso, ao final desse processo, o bioproduto de interesse precisa ser separado dos demais compostos que estão presentes na solução formada (3,7,14). As técnicas de pré-tratamento, ruptura celular e recuperação são de suma importância para a obtenção do produto.
Portanto, o objetivo deste trabalho foi avaliar diferentes pré-tratamentos na biomassa seguido de ruptura celular com dimetilsulfóxido (DMSO), visando maior recuperação de carotenoides e redução no tempo de processo.
MATERIAL E MÉTODOS
Micro-organismo
A levedura Rhodotorula mucilaginosa CCT 7688 utilizada foi previamente isolada, identificada e caracterizada por Otero et al. (13) a partir de amostras ambientais da região Sul do Rio Grande do Sul – Brasil, e depositadas na Coleção de Culturas Tropicais André Tosello (Campinas, Brasil).
Manutenção, reativação e cultivo do inóculo da cultura microbiana
A levedura foi mantida em tubos com ágar YM (Yeast-Malt extract) inclinado, sendo esse composto por 10 g L-1 de glicose, 5 g L-1 de peptona, 3 g L-1 de extrato de malte, 3 g L– 1 de extrato de levedura e 20 g L-1 de ágar bacteriológico. No processo de reativação, os tubos foram incubados a 25 °C por 48 h. Para o preparo do pré-inóculo a biomassa foi ressuspensa em 1 mL de água peptonada 0,1 % e transferida para tubos com 9 mL de caldo YM, sendo incubados por 48 h a 25 °C. O cultivo do inóculo foi feito em frascos Erlenmeyer (250 mL) contendo 90 mL de caldo YM, sob agitação de 150 rpm, 25 ºC e por 48 h (8).
Ensaios para a produção de carotenoides
Os cultivos foram feitos utilizando frascos Erlenmeyer agitados (500 mL), os quais continham 225 mL de meio agroindustrial com 70 g L-1 de melaço e 3,4 g L-1 de água de maceração de milho. O pH inicial foi 6,0, 10 % de inóculo (v v-1), 25 °C e 180 rpm em agitador orbital (8). A determinação da concentração (biomassa e carotenoides) e pH foram feitos ao final de 144 h de cultivo.
Determinação da concentração de biomassa e pH
A concentração de biomassa foi determinada pela leitura de transmitância a 620 nm em espectrofotômetro (Quimis, modelo Q998U, Brasil) e utilizando a curva padrão de biomassa (16). Para a determinação do pH, foi utilizado pHmetro de bancada (Kasvi, modelo K39-1420A, Brasil), segundo AOAC (17).
Pré-tratamento da biomassa, extração e determinação da concentração de carotenoides
A biomassa ao final dos cultivos (144 h) foi separada por centrifugação, lavada e ressuspendida em água destilada. Para os testes de extração, a biomassa foi submetida a dois diferentes pré-tratamentos: (a) secagem da biomassa por 24 h em estufa a 35 ºC seguida de congelamento em freezer à -18 ºC por 24 h; (b) secagem da biomassa por 48 h em estufa a 35 ºC seguida de congelamento em freezer à -18 ºC por 48 h. A biomassa de ambos os pré-tratamentos foi macerada com gral e pistilo e padronizada o tamanho da partícula utilizando peneira mesh 115 antes de submetidas a operação de congelamento.
Após a realização dos pré-tratamentos na biomassa, foram realizados quatro testes de extração, conforme demonstrado na Figura 1, onde os tratamentos T1 e T3 diferenciam-se de T2 e T4 pelo ciclo de ruptura da célula com DMSO. Sendo os tratamentos T1, T2 e T3 propostos por Silva et. (7) comparados com o tratamento T4 proposto por Rodrigues et al. (8), considerado o método padrão para a extração de carotenoides por leveduras carotenogênicas utilizando DMSO.
Nos tratamentos T1 e T3, em 0,05 g de biomassa acrescentou-se 2 mL de DMSO à 55 °C, homogeneizou-se por 1 min em vórtex (Biomixer QL-901, Ningbo, China) para o rompimento da célula, posteriormente adicionou-se 6 mL de acetona para a recuperação dos carotenoides e o processo foi repetido até o branqueamento das células. Os tratamentos T2 e T4, diferenciaram-se dos demais devido a homogeneização por 1 min em vórtex (Biomixer QL-901, Ningbo, China) com intervalos de 15 min até completar 1 h, posteriormente adicionou-se 6 mL de acetona para a recuperação dos carotenoides e o processo foi repetido até o branqueamento das células.
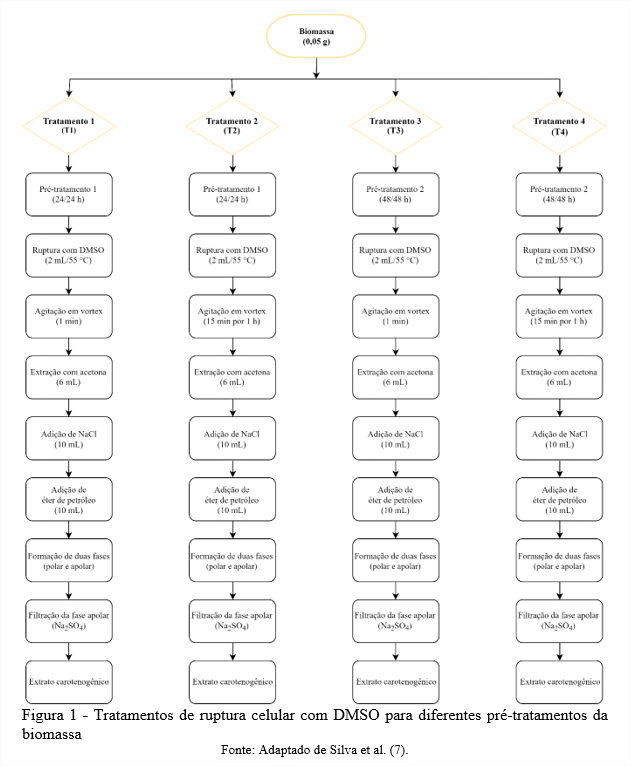
Após o processo de extração com os diferentes tratamentos de ruptura química com DMSO, o sobrenadante foi reservado e adicionado 10 mL de solução de NaCl 20 % (m v-1) e 10 mL de éter de petróleo. Após a formação de duas fases com diferentes polaridades, utilizou-se do sulfato de sódio (Na2SO4) para filtrar a fase apolar formada, obtendo-se os extratos carotenogênicos (4,8).
Segundo Goodwin (3), o β-caroteno é o carotenoide majoritário da R. mucilaginosa e tem absortividade específica de 2592 em éter de petróleo. Com este dado, a leitura de transmitância em espectrofotômetro (Quimis, modelo Q998U, Brasil) à 450 nm (10,12),determinou-se a concentração específica de carotenoides (CEC), utilizando a Equação 1 (Eq. 1), sendo V o volume em mL, A absorbância e a biomassa seca em g.. Com os resultados da CEC e concentração de biomassa, foi calculada a concentração volumétrica de carotenoides – CVC (μg L-1) (10).

Análises estatísticas
Os ensaios de extração de carotenoides foram realizados em triplicatas e os resultados submetidos à Análise de Variância (ANOVA) e as médias comparadas pelo Teste de Tukey com 95 % de confiança.
RESULTADOS E DISCUSSÃO
Baseando-se nas pesquisas feitas por Silva et al. (7) e Rodrigues et al. (8), os tratamentos feitos tinham como objetivo definir o pré-tratamento mais eficiente para fazer a ruptura celular e extração dos carotenoides produzidos pela levedura Rhodotorula mucilaginosa CCT 7688. Os dados da Tabela 1 mostram que não houve diferença significativa entre os tratamentos T2 e T4 – os quais foram submetidos a diferentes pré-tratamentos da biomassa (tempo de secagem e congelamento), mas ao mesmo processo de extração, utilizando ciclos de ruptura celular de 15 min até 1 h.

A melhor resposta aos tratamentos aplicados foi encontrada em T1, com CEC de 272,81 μg g-1, sendo diferente significativamente dos demais tratamentos (p<0,05) e 36,4 % mais eficiente na extração de carotenoides do que o tratamento T4, metodologia considerada padrão pela literatura.
Michelon et al. (15) avaliaram para a levedura Phaffia rhodozyma NRRL Y-17268, a utilização de diferentes técnicas de ruptura celular, dentre elas a utilização de DMSO – o mesmo utilizado por Rodrigues et al. (8) – com a aplicação de congelamento (−18 °C por 48 h). O estudo apontou que o pré-tratamento (congelamento da biomassa) aumentou em 16 % a concentração de carotenoides quando comparado ao controle, o qual foi feita a extração sem a submissão ao processo de ruptura celular.
Os resultados obtidos foram superiores aos encontrados por Rodrigues et al. (8) com a mesma levedura e o mesmo meio agroindustrial utilizando o pré-tratamento de secagem por 48 h e congelamento por 48h e ruptura com DMSO, sendo a melhor concentração de carotenoides volumétricos de 1248,5 μg L-1. Machado et al. (18) cultivaram R. toruloides URM 7406 em meio YM para produção de carotenoides e com o mesmo processo de ruptura e pré-tratamento de biomassa descritos acima e atingiu 1333,11 µg L–1.
Fonseca et al. (14) também utilizaram pré-tratamentos e método de ruptura celular com DMSO nas células da levedura Phaffia rhodozyma NRRL Y-17268. Sendo constatado que a quantidade de água fora da célula, como também a água livre, tem influência quanto aos processos de ruptura celular e de extração de carotenoides. Assim, o uso dos pré-tratamentos na biomassa da levedura, diminuíram a quantidade de água livre, fazendo com que fosse recuperada maior concentração de carotenoides (375,7±10,5 μg g-1).
Silva et al. (7) avaliaram diferentes pré-tratamentos térmicos da biomassa para a extração de carotenoides produzidos por Phaffia rhodozyma NRRL Y-17268: liofilização, secagem e congelamento, além do tempo de ruptura química das células com o uso de DMSO: agitação por 1 min em vórtex e agitação (método 1) e a cada 15 min por 1 h (método 2). O pré-tratamento da biomassa com secagem por 24 h e congelamento por 24 h, associado ao processo químico de ruptura com agitação de 1 min se mostrou eficaz na extração dos carotenoides, obtendo-se rendimento de até 50 % quando comparado ao método 2.
Portanto, através da literatura consultada e os tratamentos aplicados neste estudo, é possível inferir que os pré-tratamento na biomassa para a levedura R. mucilaginosa são necessários, sendo que dentre os avaliados, o mais eficiente foi a secagem por 24 h seguido de congelamento por 24 h, alinhado ao processo de ruptura celular com DMSO descrito em T1 (Figura 1). Desta forma, utilizando este procedimento foi possível reduzir o tempo de processo em 48 h quando comparado com a metodologia comumente utilizada e descrita na literatura.
CONCLUSÃO
O método mais adequado para o pré-tratamento e extração de carotenoides, com 95 % de confiança, é o tratamento 1 (T1), proposto por Silva et al. (7). Esse demonstrou ser a combinação mais eficiente de pré-tratamento e extração, apresentando os melhores rendimentos (272,81 μg g-1), reduzindo o tempo de processo em 48 h, além de ser 36,4 % mais eficiente quando comparado a metodologia padrão utilizada de acordo com a literatura.
AGRADECIMENTOS
Os autores agradecem ao apoio da Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Código de Financiamento 001; ao CNPq pela bolsa de iniciação científica e bolsas de produtividade em pesquisa; a FAPERGS e a Universidade Federal do Rio Grande (FURG).
REFERÊNCIAS
1. Chandra P, Sharma RK, Arora DS. Antioxidant compounds from microbial sources: A review. Food Res Int. 2020;129:108849.
2. Rao AR, Baskaran V, Sarada R, Ravishankar GA. In vivo bioavailability and antioxidant activity of carotenoids from microalgal biomass — A repeated dose study. Food Res Int. 2013;54(1):711–7.
3. Goodwin TW. The Biochemistry of the Carotenoids. 2. ed. Vol. 1. Dordrecht: Springer Netherlands; 1980.
4. Chociai MB, Machado IMP, Fontana JD, Chociai JG, Busato SB, Bonfim TMB. Cultivo da levedura Phaffia rhodozyma (Xanthophyllomyces dendrorhous) em processo descontínuo alimentado para produção de astaxantina. Rev Bras Ciênc Solo. [Internet]. 2002 [acesso em 25 Out 2022];38(4). Disponível em: http://www.scielo.br/scielo.php?script=sci_arttext&pid=S1516-93322002000400008&lng=pt&nrm=iso&tlng=pt
5. Prescendo Júnior D, Silva PGP, Burkert JFM, Santos LO. Aprimoramento da etapa downstream para a extração de carotenoides produzidos pela levedura Phaffia rhodozyma. Rev Bras Agrotecnologia. 2021;11(2):71–6.
6. Rodriguez-Amaya DB. Update on natural food pigments – A mini-review on carotenoids, anthocyanins, and betalains. Food Res Int. 2019;124:200–5.
7. Silva PGP, Prescendo Júnior D, Burkert JFM, Santos LO. Carotenoid extraction from Phaffia rhodozyma biomass: downstream strategies and economic evaluation of energy. Braz J Chem Eng. [Internet]. 2022 [acesso em 26 Out 2022]; Disponível em: https://link.springer.com/10.1007/s43153-022-00225-7
8. Rodrigues TVD, Amore TD, Teixeira EC, Burkert JFM. Carotenoid production by Rhodotorula mucilaginosa in batch and fed-batch fermentation using agroindustrial byproducts. Food Technol Biotechnol. 2019;57(3):388–98.
9. Valduga E, Tatsch PO, Tiggemann L, Treichel H, Toniazzo G, Zeni J, et al. Produção de carotenoides: microrganismos como fonte de pigmentos naturais. Quím Nova. 2009;32:2429-2436.
10. Cipolatti EP, Remedi RD, Sá CS, Rodrigues AB, Ramos JMG, Burkert CAV, et al. Use of agroindustrial byproducts as substrate for production of carotenoids with antioxidant potential by wild yeasts. Biocatal Agric Biotechnol. 2019;20:101208.
11. Rios DAS, Borba TM, Burkert JFM, Kalil SJ. Rice parboiling wastewater in the maximization of carotenoids bioproduction by Phaffia rhodozyma. Ciênc Agrotec. 2015;39:401–10.
12. Silva CM, Borba TM, Burkert CAV, Burkert JFM. Carotenoid production by Phaffia rhodozyma using raw glycerol as an additional carbon source. Int J Food Eng. [Internet]. 2012 [acesso em 23 Out 2022];8(4). Disponível em: https://www.degruyter.com/document/doi/10.1515/1556-3758.2843/html
13. Otero DM, Bulsing BA, Huerta KM, Rosa CA, Zambiazi RC, Burkert CAV, et al. Carotenoid-producing yeasts in the brazilian biodiversity: isolation, identification and cultivation in agroindustrial waste. Braz J Chem Eng. 2019;36(1):117–29.
14. Fonseca RAS, Rafael RS, Kalil SJ, Burkert CAV, Burkert JFM. Different cell disruption methods for astaxanthin recovery by Phaffia rhodozyma. Afr J Biotechnol. 2011;10:1165–71.
15. Michelon M, Borba TM, Rafael RS, Burkert CAV, Burkert JFM. Extraction of carotenoids from Phaffia rhodozyma: A comparison between different techniques of cell disruption. Food Sci Biotechnol. 2012;21(1):1–8.
16. Choi MH, Park YH. Production of yeast biomass using waste Chinese cabbage. Biomass Bioenergy. 2003;25(2):221–6.
17. Association of Official Analytical Chemists (AOAC). Official Methods of Analysis. 17. ed. Arlington, Va; 2000.
18. Machado WRM, Silva LG, Vanzela ESL, Bianchi VLD. Production of carotenoids by Rhodotorula toruloides isolated from brazilian tropical savannah. Int. Food Res. J. 2019;26:1259–1267.
Capítulo de livro publicado no Congresso Brasileiro de Química dos Produtos Naturais. Para acessa-lo clique aqui.
Este trabalho foi escrito por:
Juliana Ayres de Araújo ; Carlos Henrique Milagres Ribeiro *; Teresa Drummond Correia ; Alex Oliveira Botelho ; Maria Carolina Marinho Nicolau ; Stephanie Regina de Jesus Souza
*Autor correspondente – Email: [email protected]
Resumo: Diante das perdas pós-colheita provenientes do surgimento de doenças, a podridão parda se destaca como a mais significativa para a cultura do pêssego [Prunus persica (L.) Batsch], fazendo-se necessário seu controle, que geralmente é realizado via aplicação de produtos químicos. Atualmente, busca-se por novos meios de controle visando uma maior sustentabilidade. Assim, este trabalho objetivou avaliar o efeito do óleo essencial de cravo-da-índia (Syzygium aromaticum) sobre o desenvolvimento do fungo Monilinia fructicola, in vivo, buscando uma alternativa a aplicação de produtos químicos no controle pós-colheita da podridão parda. O experimento foi conduzido em delineamento inteiramente casualizado (DIC), com seis tratamentos e seis repetições, em diferentes concentrações do óleo de cravo-da-índia em água destilada. Os tratamentos consistiram de: 1- Ausência da aplicação do óleo essencial de cravo (testemunha), 2- 0,25%, 3- 0,5%, 4- 0,75%, 5- 1,0%, 6- 1,25%. Realizou-se a inoculação do fungo Monilinia fructicola por borrifação, e 15 horas depois, aplicou-se os tratamentos, imergindo os frutos por 30 segundos. As variáveis analisadas, após 7 dias, foram a incidência e severidade da doença. Os resultados demonstraram que o óleo de cravo não foi eficiente no controle de Monilinia fructicola.
Palavras–chave: podridão parda, controle alternativo, Prunus persica (L.) Batsch.
Abstract: Towards of the post-harvest losses due to the appearance of diseases, brown rot stands out as the most significant for the peach [Prunus persica (L.) Batsch] production, requiring its control, which is usually done by application of chemicals products. Currently, it has been sought for new ways of disease control aiming at greater sustainability. Thereby, the objective of this work was to evaluate the effect of clove essential oil (Syzygium aromaticum) on the development of the fungus Monilinia fructicola, in vivo, searching for an alternative to the application of chemicals in the post-harvest control of brown rot. The experiment was conducted in a completely randomized design (CRD), with six treatments and six replicates at different concentrations of clove oil in distilled water. The treatments consisted of: 1- No application of clove essential oil (control), 2- 0,25%, 3- 0,5%, 4- 0,75%, 5- 1,0%, 6- 1, 25%. The fungus Monilinia fructicola was inoculated by spraying, and 15 hours later, the treatments were applied, immersing the fruits for 30 seconds. The variables analyzed after 7 days were the incidence and severity of the disease. The results demonstrated that clove oil was not efficient in the control of Monilinia fructicola.
Key Word: Brot; alternative control; Prunus persica (L.) Batsch.
INTRODUÇÃO
O pêssego [Prunus persica (L.) Batsch] é originário da China e rapidamente se tornou uma das espécies de clima temperado mais cultivadas em diversas regiões (1, 2). É considerada a oitava fruta mais cultivada mundialmente, sendo a China o principal produtor. O Brasil se encontra como o décimo na produção mundial (3).
No Brasil, a maioria da sua produção se concentra nas regiões Sul e Sudeste, que possuem condições climáticas favoráveis para seu desenvolvimento (4). Os Estados que detém o maior número de produtores são Rio Grande do Sul, São Paulo, Santa Catarina, Minas Gerais, Paraná (5). Embora o cultivo de pêssego esteja se expandindo para outros Estados brasileiros onde há temperaturas mais elevadas, a área de sua produção reduziu 10,85% entre os anos de 2010 e 2013, no entanto, houve elevação de sua produtividade em 9,8% no mesmo intervalo de tempo (6). Esse aumento se deve a aplicação de tecnologias e melhorias de manejo que visam a adaptabilidade do pêssego a diferentes regiões brasileiras.
A produção do pêssego pode ser encaminhada tanto para o mercado de processamento industrial, quanto para o consumo in natura. A demanda da fruta é baixa no país, mas ainda assim a produção nacional não consegue supri-la, necessitando de importações (7) e demonstrando que seu cultivo é um mercado promissor a ser explorado.
Para se estabelecer nesse mercado rentável é preciso lidar com um dos grandes entraves encontrados na persicultura, as perdas decorrentes da pós-colheita. Como uma das suas principais causas está o ataque de agentes patogênicos causadores de podridões, como a podridão-parda, provocada pelo fungo Monilinia fructicola (8). Essa é a principal doença da cultura (9) e o método de controle mais utilizado é através da aplicação de fungicidas desde a floração até a pré-colheita (10). Na fase de pós-colheita, há restrição legislativa quanto ao uso de produtos químicos, visto que esses são propícios a resíduos tóxicos nos frutos (11).
A utilização excessiva de produtos químicos pode ocasionar a resistência do patógeno, e, além disso, cada vez mais os consumidores questionam sua aplicação, devido à busca por alimentos saudáveis, livres de resíduos tóxicos (12).
Trabalhos e pesquisas sobre métodos alternativos de controle têm sido elaborados para que essas questões sejam resolvidas, e uma das opções estudadas é o uso de óleos essenciais, que têm demonstrado potencial para a inibição da atividade fúngica, pois apresentam um ou mais compostos responsáveis pelo(s) efeito(s) causado(s) em suas diferentes aplicações, onde um composto majoritário geralmente atua (13).
Atualmente tem se observado trabalhos positivos com a utilização do óleo essencial de cravo-da-índia (Syzygium aromaticum) no controle antimicrobiano (14, 15). Porém, não existem trabalhos que comprovem a afetividade do óleo essencial de cravo-da-índia no desenvolvimento de fungo em pós-colheita do pêssego.
Diante disso, o objetivo desse trabalho foi avaliar o efeito do óleo essencial de cravo-da-índia sobre o desenvolvimento do fungo Monilinia fructicola, in vivo, como uma alternativa a aplicação de produtos químicos no controle da podridão parda.
MATERIAL E MÉTODOS
O experimento foi conduzido no Laboratório de Fitossanidade, situado no Núcleo de Agricultura do Instituto Federal do Sudeste de Minas Gerais – Campus Barbacena.
Isolamento e cultivo de M. Fructicola
O fungo M. fructicola foi isolado a partir de frutos mumificados oriundos do pomar de pêssego do Instituto Federal Sudeste de Minas Gerais – Campus Barbacena, realizando-se a identificação do fungo no laboratório de fitossanidade.
Os procedimentos foram executados na câmara de fluxo laminar, atendo-se aos devidos cuidados quanto à assepsia necessária, de acordo com a metodologia descrita para isolamento de fungos fitopatogênicos (16). Os isolados foram cultivados, para seu desenvolvimento, em meio de cultura BDA (batata-dextrose-ágar) e dispostos em incubadora B.O.D (Demanda Bioquímica de Oxigênio) com temperatura de 25º C e fotoperíodo de 12 h. Após 10 dias, realizou-se a repicagem micelial para a obtenção da cultura fúngica pura, até atingir toda a superfície da placa. A cultura pura do fungo continuou armazenada na incubadora por 23 dias.
Obtenção dos frutos e inoculação de M. fructicola
A aquisição dos pêssegos da cv. Rubimel foi feita através de um pomar comercial da Chácara da Mantiqueira, uma empresa produtora de frutas selecionadas, localizada na BR 040, KM 705 – Galego – Barbacena – Minas Gerais.
Os frutos foram colhidos manualmente no início do mês de novembro, época que caracteriza o início da maturação da cultivar selecionada. Considerou-se a uniformidade de tamanho, cor e ausência de qualquer tipo de injúria ou sintomas de doenças.
A suspensão dos esporos ocorreu via adição de aproximadamente 10 mL de água destilada na cultura fúngica. Utilizando uma alça Drigaslki, raspou-se os conídios e micélio, que foram submetidos a filtragem feita no algodão. A solução foi colocada no agitador Vortex por 1 minuto, sob severa agitação. A concentração de inóculo presente na suspensão foi calibrada na câmara de Neubauer para 105 esporos/mL.
Foi realizado somente um ferimento em cada fruto de aproximadamente 5 mm de profundidade e 0,27 mm de diâmetro, feito com uma agulha hipodérmica previamente esterilizada e, em seguida, a área foi demarcada ao seu redor. A inoculação foi feita através de borrifação sobre a área demarcada com o ferimento, até o ponto de escorrimento.
Os frutos foram, então, colocados em bandejas plásticas para o acondicionamento em câmara úmida, que foram cobertas com papel filme PVC por aproximadamente 15 horas para garantia da ocorrência da infecção, não havendo ainda a aplicação dos tratamentos.
Tratamento com óleo de cravo-da-índia (Syzygium aromaticum)
O óleo de cravo-da-índia foi adquirido no comércio local de Barbacena-MG. Antes da aplicação do óleo de cravo-da-índia, os frutos foram pesados para obtenção da homogeneidade de peso dos frutos usados no experimento e inoculados.
O experimento constituiu-se por 6 tratamentos, dispostos na Tabela 1.

Todos as concentrações de óleo de cravo-da-índia foram diluídas em água destilada esterilizada, totalizando 300 mL de solução.
Para aplicação dos tratamentos, os frutos foram imersos na solução final, que foi colocada em um béquer de 1000 mL, sendo um béquer para cada tratamento, durante 30 segundos, sob agitação manual da solução. Os frutos foram retirados da solução e colocados para secagem em papel toalha em temperatura ambiente, e em seguida foram distribuídos individualmente em potes de plástico forrados com papel toalha, que foram colocados de modo aleatório em 6 bandejas plásticas, com 6 repetições em cada uma.
As bandejas foram mantidas em temperatura ambiente.
Parâmetros analisados
Para os parâmetros, considerou-se a avaliação da incidência e a severidade da doença. A incidência foi analisada considerando a presença ou ausência do patógeno. Para a severidade da doença, foram atribuídas notas de 1 a 5 baseando-se na escala visual (17), sendo consideradas as seguintes notas de acordo com a superfície com lesão disposto na Tabela 2.

As avaliações foram realizadas após 7 dias da inoculação do fungo. Para tal, observou-se visualmente a presença ou ausência de sintomas e sinais da doença. Considerou-se a presença de micélios com aspecto cotonoso característico para o fungo, bem como a mancha parda, caracterizada por podridão aquosa.
Ressaltando que todas as avaliações foram feitas pelo mesmo avaliador, para evitar resultados tendenciosos.
O delineamento experimental adotado foi inteiramente casualizado (DIC), com 6 tratamentos e 6 repetições, constituindo-se de 36 parcelas experimentais, sendo a unidade experimental constituída de 1 pêssego
Os valores obtidos para os parâmetros analisados, foram submetidos a análise de variância (ANOVA), sendo feita a curva de regressão (p>0,05%) e as médias foram comparadas pelo teste Scott-Knott a 5% de probabilidade. Para o auxílio dos cálculos estatísticos utilizou-se o programa SISVAR (18).
RESULTADOS E DISCUSSÃO
Após os sete dias da inoculação, realizou-se a avaliação dos resultados, cujas análises de variância podem ser observadas na Tabela 3, que demonstraram a existência de significância para a variável incidência (p>0,05), e ausência de significância para a variável severidade (p>0,05). Ambas as médias podem ser observadas nas Figuras 1 e 2, respectivamente.

Ao analisar os frutos, verificando a presença de sinais e sintomas da podridão parda, a testemunha apresentou melhor resultado com relação a incidência (Figura 1), quando comparada aos demais tratamentos que utilizaram o óleo essencial de cravo-da-índia em concentrações diferentes, sendo sua incidência completamente zerada.
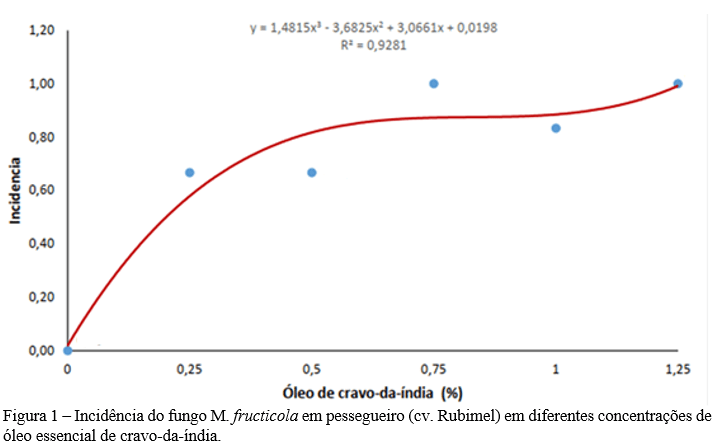
Embora não tenham sido avaliadas as características físico-químicas dos
pêssegos, nas condições experimentais, é importante mencionar que foi possível observar que grande parte dos frutos apresentaram manchas marrons e depressão no epicarpo (Figura 2), logo após a imersão dos mesmos no óleo diluído, tornando-se inviáveis para a comercialização.

Assim, acredita-se que o óleo de cravo-da-índia reagiu com os frutos
ocasionando toxidez, visto que qualquer substância pode ser vista como um agente tóxico, variando de acordo com as condições de exposição, dose aplicada
ou absorvida, tempo e frequência de exposição e modo de aplicação (19).
Alguns frutos foram totalmente recobertos por essas manchas, outros
parcialmente, onde foi possível notar a presença de manchas circulares, e o restante
não sofreu alteração visual. Ressaltando que essas manchas não se caracterizam
como manchas pardas típicas da podridão parda, não sendo aquosas.
Quanto ao surgimento de depressões, essas se concentraram nas regiões
onde o fruto apresentou manchas marrons circulares, sendo notável que as mesmas
se intensificaram durante o intervalo entre seu surgimento após a imersão do fruto nos
tratamentos e o dia da avaliação.
Mesmo não ocorrendo diferença para o parâmetro analisado de severidade, observa-se que houve uma diferença entre as médias da severidade do fungo presente na Figura 3.
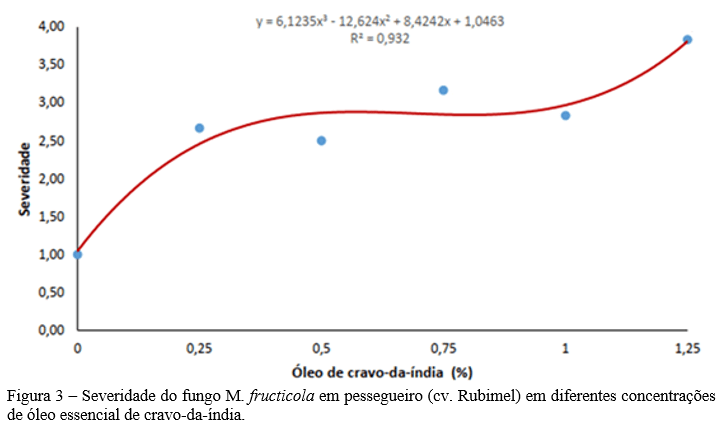
Além disso, nos mesmos locais referentes à alteração na coloração do epicarpo diante dos tratamentos, observou-se ainda, modificação na coloração do mesocarpo nos frutos (Figura 4). As modificações mencionadas não apresentaram distinção visual de progressão entre as diferentes concentrações do óleo.

Alguns autores nos remetem a potencialidade do óleo de cravo-da-índia no
controle de patógenos (20), que ao testar a concentração de 25% de extrato hidroalcoólico de cravo-da-índia, obteve inibição total de Fusarium pallidoroseum até o sétimo dia de incubação do meloeiro.
No experimento analisando o efeito de óleo essencial de cravo-da-índia em goiabeira (21), observou-se que na concentração de 10% do óleo, in vitro, houve inibição completa do crescimento micelial. Entretanto, não foi observado controle do micélio, ao se testar a concentração de 2,5%, in vivo.
O eugenol, principal componente do óleo de cravo-da-índia, é um fenol volátil, sendo mencionado como o responsável pelos efeitos fungicidas do composto (22). Com relação aos resultados negativos encontrados no presente estudo, a característica de volatilidade pode ser apontada como a possível explicação, visto que, durante o experimento, após a aplicação dos tratamentos contendo o óleo de cravo-da-índia notou-se no ambiente ao decorrer dos primeiros dias, a presença de um forte aroma.
O resultado deste trabalho, infere que apesar do óleo de cravo-da-índia ser descrito por diversos pesquisadores como uma opção no controle alternativo à variadas doenças, diante das condições e metodologia que este experimento foi desenvolvido, o mesmo atuou de modo antagônico. O óleo essencial de cravo-da-índia, além de não se apresentar como eficiente no controle da incidência e severidade patogênica, demonstrou fitotoxicidade, causando lesões no epicarpo do fruto, favorecendo as condições para que o patógeno se desenvolvesse.
No trabalho utilizando o tratamento com o óleo de cravo-da-índia na concentração de 100 µL/100 mL de água via perfuração de orifício seguida de inoculação de Colletotrichum musae após 12 horas de incubação (23), constataram que as cascas da maioria das bananas ficaram completamente escuras na maioria das repetições, provavelmente devido a alguma atividade fitotóxica desse óleo na concentração que foi testada. Já em estudo realizado com óleos essencial do cravo-da-índia, objetivando a qualidade de feijão- fava (24), os autores verificaram que na concentração de 2 mL. L-1, atuou de modo prejudicial a planta, reduzindo a qualidade fisiológica das sementes.
O não desenvolvimento da doença na testemunha pode ser justificada através da investigação de tratos culturais no pomar de origem dos frutos, onde verificou-se a aplicação do produto químico inorgânico Kumulus® DF, que atua contra o desenvolvimento de agentes patogênicos externos.
No experimento visando alternativas do controle pós-colheita da podridão-parda e da podridão-mole em frutos de pessegueiro (25), ao aplicarem o óleo de cravo-da-índia na concentração de 0,01% no tratamento pós-colheita do pêssego, não obteve redução da incidência e nem da severidade da podridão-parda, sendo suas médias maiores do que as apresentadas pela testemunha.
Outro fator passível de contribuição para que o óleo de cravo-da-índia não controlasse o fungo M. fructicola, deve-se a suas características de instabilidade diante de fatores como a presença de calor, ar, luz e umidade, o que pode gerar a rápida evaporação de seus componentes (23).
Em reação aos efeitos preventivos e curativos, é provável que o óleo de cravo-da-índia se sobressaia melhor diante do efeito preventivo, em meio de cultura antes da inoculação, do que perante efeito curativo, com a imersão dos frutos e contaminação provenientes do campo (26).
Os trabalhos realizados em âmbito de controle alternativo utilizando óleos
essenciais in vivo ainda são escassos, havendo ampla diversidade quanto a
metodologia aplicada, o que torna difícil a formação de uma tomada de decisão
concisa.
Sua eficiência tende a variar de acordo com o óleo essencial escolhido,
metodologia aplicada, patógeno alvo e susceptibilidade da variedade que receberá o tratamento. E, mesmo que haja o controle da doença, faz-se necessária uma metodologia que mantenha os aspectos físico-químicos da planta.
CONCLUSÕES
Os resultados apresentados mostram que o óleo essencial de cravo-da-índia (Syzygium aromaticum) não foi efetivo no controle de Monilinia fructicola na pós-colheita.
Além disso, o efeito causado pelo óleo de cravo-da-índia no epicarpo e
mesocarpo tornaram os pêssegos inviáveis para comercialização, visto que os
consumidores tendem a comprar pelo aspecto visual do fruto.
Diante do exposto trabalho, considerando o método utilizado, o óleo de cravo-da-índia não se apresenta como uma alternativa a aplicação de produtos químicos no controle pós-colheita da podridão parda.
REFERÊNCIAS
- De Souza AM., Ayub RA, Pinto CA. Adaptabilidade de cultivares de pessegueiro na região dos Campos Gerais–Paraná. Rev. em Agronegócio e Meio Ambient. 2022;15; 1-14
- RASEIRA MCB. Pêssego cultivar BRSbrs libra. Rev. Bras Fruti. 2010; 32:961-1296.
- Fao. Food and agriculture organization of united nations – FAO. Faostat. Roma: FAO, [Internet]. 2021. [acesso em 2020 14 dez]. Disponível Disponível em: https://www.fao.org/faostat/es/#data/QCL .
- D’avila RF, Zambiazi RC, Sá OS, Toralles RP. Atividade de ßglucosidases em extrato enzimático obtido de amêndoas de pêssego. . Rev. Bras Fruti.2015;37:541-549.
- Faostat (Food and Agriculture Organization of the United Nations Statistics Division). On-line database. [Internet]. 2017 [acesso em 2022 14 dez]. Disponível em: http://www.fao.org/faostat/. Rome, Italy.
- Seplan . Rio grande do sul. Secretaria do Planejamento e Desenvolvimento Regional (Seplan). Atlas Socioeconômico do Rio Grande do Sul: economia: pêssego e banana – 2020. [Internet]. 2021. [acesso em 2020 14 dez]. Disponível em: https://atlassocioeconomico.rs.gov.br/pessego-e-banana.
- Madail JCM, Raseira MCB. Aspectos da produção e mercado do pêssego no Brasil. [Internet]. Pelotas: Embrapa Clima Temperado; Circular Técnica 80; 2008. [acesso em: 15 dez 2022]. Disponível em: http://www.agencia.cnptia.embrapa.br/Repositorio/Circular_80_000gihms14i02wx5ok05vadr1g2w5svd.pdf.
- Sautter CK, Brackmann A, Anese RDO, Weber A, Rizzatti MR, Pavanello EP. Controle da podridão-parda e características físico-químicas de pêssegos ‘Magnum’ submetidos a tratamentos pós-colheita com elicitores abióticos. Rev Ceres. 2011; 58:172-177.
- Moreira LM, May-de mio LL. Controle da podridão parda do pessegueiro com fungicidas e fosfitos avaliados em pré e pós-colheita. Ciên agrotec. 2009; 33: 405-411.
- May-de mio LL, Garrido L, Ueno B. Doenças de fruteiras de caroço. In: MONTEIRO LB. Fruteiras de caroço: uma visão ecológica. 2004.
- Moreira LM. Alternativas de controle integrado da podridão parda do pessegueiro. [Tese]. Curitiba; Universidade Federal do Paraná; 2005.
- Franca, D. As técnicas de controle biológico projetam um futuro promissor e lucrativo para a agricultura: As técnicas inovadoras do conceito de Controle Biológico têm se revelado cada vez mais promissoras. Inova. & Desen: A Rev da FACEPE. 2020; 1: 11-13.
- Lorenzetti ER. Controle de doenças do morangueiro com óleos essenciais e Trichoderma spp. [Tese]. Lavras: Universidade Federal de Lavras; 2012.
- Tarek N, Hassan H, Abdelghani SMM, Radwan, IA, Hammouda O, El-gendy AO. Comparative chemical and antimicrobial study of nine essential oils obtained from medicinal plants growing in Egypt. Beni-suef University Jour Of Basic and Applied Scien.2014;3:149-156.
- Almeida AC, Oliveira L, Paulo PD, Martins ER, Souza RM, Figueiredo LS, Santos CA, Fonseca HC. Potencial antimicrobiano dos óleos essenciais de cravo-da-índia (Syzygium aromaticum L.) e alfavacão (Ocimum gratissimum L.) em carne moída de ovinos contaminada experimentalmente com Staphylococcus aureus. Rev. Bras de Ciên Vet. 2013; 20:248-251.
- Alfenas AC. Isolamento de fungos fitopatogênicos. In: MAFIA, R.G. Métodos em Fitopatologia. 1. UFV; 2016.
- Wagner AJR, Raseira MCB, Fortes JF, Pierobom CR, Silva JB. Non-Correlation of Flower and Fruit Resistance to Brown Rot (Monilinia fructicola (Wint.) Honey) Among 27 Peach Cultivars and Selections. Jour of the Ameri Pomo Socie. 2005; 59: 148-152.
- Ferreira DF. Sisvar: a computer statistical analysis system. Ciên Agrotec. 2011; 35:1039-1042.
- Affonso RS, Rennó MN, Slana GB, Franca TC. Aspectos Químicos e Biológicos do Óleo Essencial de Cravo da Índia. Rev. Virt Quí. 2012; 4:146-161.
- Coutinho IBL, Gagliardi PR, Ootani MA. Atividades de extratos vegetais no controle de Fusarium pallidoroseum (Cooke) Sacc em meloeiro. Essentia. 2015; 16:40-61.
- Rozwalka LC. Controle alternativo da antracnose em frutos de goiabeira, em laboratório. [dissertação]. Curitiba:Universidade Federal do Paraná; 2003.
- Pereira AA. Efeito inibitório de óleos essenciais sobre o crescimento de bactérias e fungos. [dissertação]. Lavras: Universidade Federal de Lavras; 2006.
- Oliveira ES, Viana FMP, Martins MVV. Alternativas a fungicidas sintéticos no controle da antracnose da banana. Sum phyto. 2016; 42: 340-350.
- GOMES RSS, Nunes MC, Nascimento LC, Souza JO, Porcino MM. Eficiência de óleos essenciais na qualidade sanitária e fisiológica em sementes de feijão-fava (Phaseolus lunatus L.). Revi. Brasi Plant Medi. 2016; 18: 279-287.
- Carvalho VL, Cunha RLD, Chalfun NNJ, Moura PHA.Alternativas de controle pós-colheita da podridão-parda e da podridão-mole em frutos de pessegueiro. Revi. Bras Fruti. 2009; 31:78-83.
- Fontana DC, Kulczynski SM, Trevisan R, Schmit D, Caron BO, Pinheiro MVM, Diel MI. Uso de extratos vegetais no controle alternativo da podridão parda do pessegueiro. Revi cult saber. 2017; 10:148-165.